细胞中的能量
能量概述
酶的探究历程
这段关于“酶”的探究历程,生动地展示了科学认知是如何通过观察、实验、争论和技术突破而不断演进的。以下是根据你提供的纲要进行的详细展开:
早期探索,从物理消化到化学消化的转折:
- 斯帕兰札尼的实验(1773 年):在 18 世纪之前,人们普遍认为胃只能通过物理摩擦来磨碎食物。斯帕兰札尼将肉块放入带孔的小金属笼内让鹰吞下,这一设计的精妙之处在于笼子能排除胃物理摩擦的影响,但允许胃液进入。肉块的消失证明了胃液中存在某种能分解有机物的化学物质。
- 施旺发现胃蛋白酶(1835 年):德国科学家施旺在研究胃腺分泌物时,发现将其与盐酸混合后,对肉类的分解能力远超盐酸单干。他成功提取出了这种有效成分,并命名为胃蛋白酶,这是人类历史上第一次从动物体内分离出酶。
核心概念:细胞代谢离不开酶:
- 细胞代谢定义:细胞内部每时每刻都在进行的成千上万种化学反应统称为细胞代谢,它是生命活动的基础。
- 酶的必要性:许多化学反应在体外需要高温、高压、强酸或强碱等剧烈条件,但细胞内环境温和且稳定(常温、常压、pH 接近中性)。在这种环境下,只有靠酶的催化作用,代谢才能高效、有序地进行。
- 案例:过氧化氢(H_2O_2)的分解:H_2O_2 是代谢产生的有害废物。细胞内的过氧化氢酶能极快地将其分解为水和氧气,单分子催化速率可达每秒 4000 万个分子。相比无机催化剂(如 Fe^{3+}),酶的催化效率高出 10^7 \sim 10^{13} 倍,本质在于酶能显著降低反应的活化能。
酶本质的深度探索历程:
酿酒中的争论(19 世纪):
巴斯德(1857 年):认为发酵是活酵母细胞代谢的结果,没有活细胞就不能发酵。
李比希:认为发酵只是纯化学反应,只需要酵母细胞死亡裂解后释放的某些物质参与即可。
毕希纳与“酿酶”(1897 年):毕希纳用石英砂研磨酵母细胞并加压过滤,发现无细胞提取液也能使葡萄糖产生酒精。这证明了引起发酵的是细胞内的物质(他称为酿酶),而非细胞生命活动本身,从而终结了上述争论。
萨姆纳证明酶是蛋白质(1926 年):在很长一段时间内,酶被认为是不解之谜。萨姆纳坚信酶是蛋白质,历时 9 年从刀豆中提取出脲酶结晶,并证明其本质。随后,胃蛋白酶、胰蛋白酶等结晶的获得确认了“绝大多数酶是蛋白质”这一共识。
核酶的发现(20 世纪 80 年代):切赫和奥尔特曼发现少数 RNA 也具有催化功能(核酶),这一发现突破了酶只能是蛋白质的传统观念,开辟了生化研究的新领域。
讨论与总结:科学方法论。
- 观点评述:巴斯德强调了生物体与代谢的联系,李比希关注化学本质;虽然各具局限性,但他们的争论聚焦了研究目标。
- 争论的意义:科学争论能促使研究者不断设计实验去证伪或证实,推动认知走向本质。
- 毕希纳的贡献:证明了酶可以脱离细胞在体外发挥催化作用。
- 萨姆纳的精神:体现了不畏艰辛、持之以恒、实事求是的科学态度。
- 酶的完整定义:酶是活细胞产生的具有催化作用的有机物,其中绝大多数是蛋白质,少数是 RNA。
酶的探究实验
这三个实验是高中生物学中关于酶的特性及其作用原理的核心探究活动。以下是根据提供的资料进行的详细展开讲解:
比较过氧化氢在不同条件下的分解:该实验旨在通过观察过氧化氢(H_2O_2)在不同条件下的分解速率,了解酶的作用原理及高效性。
- 实验原理:H_2O_2 在常温下分解极慢,但在加热、无机催化剂(如 Fe^{3+})或新鲜肝脏研磨液(含过氧化氢酶)的作用下会加速分解产生 O_2。可以通过气泡产生的多少和带火星的卫生香复燃的剧烈程度来判断反应速率。
- 实验步骤与预期现象:
- 1 号试管(对照组):常温处理,几乎无气泡产生,卫生香不复燃。
- 2 号试管(受热):90℃ 水浴加热,产生少量气泡,卫生香不复燃。这说明加热能为反应提供能量,提高反应速率。
- 3 号试管(无机催化剂):滴加 2 滴 3.5\% FeCl_3 溶液,产生较多气泡,卫生香复燃但不明显。
- 4 号试管(过氧化氢酶):滴加 2 滴 20\% 新鲜肝脏研磨液,产生大量气泡,卫生香迅速复燃。
- 实验结论:
- 酶具有催化作用。
- 高效性:酶的催化效率远高于无机催化剂(约 10^7 \sim 10^{13} 倍),其本质是酶降低化学反应活化能的作用更显著。
- 注意事项:必须使用新鲜的肝脏研磨液,否则过氧化氢酶会被微生物分解或因放置时间过长而失活。
淀粉酶对淀粉和蔗糖的水解作用:该实验用于证明酶具有专一性,即一种酶只能催化一种或一类化学反应。
- 实验原理:淀粉和蔗糖是非还原糖,但它们在相应酶的作用下水解产生的麦芽糖、葡萄糖、果糖等属于还原糖。还原糖与斐林试剂在水浴加热条件下会产生砖红色沉淀。
- 实验设计与步骤:
- 1 号试管:注入 2\text{ mL} 淀粉溶液 + 2\text{ mL} 淀粉酶溶液。
- 2 号试管:注入 2\text{ mL} 蔗糖溶液 + 2\text{ mL} 淀粉酶溶液。
- 将两管放入 60℃(或 37℃)温水中保温 5 分钟,然后加入斐林试剂并水浴加热。
- 实验现象:1 号试管出现砖红色沉淀,2 号试管保持蓝色(斐林试剂本色)。
- 结论:淀粉酶只能催化淀粉水解,不能催化蔗糖水解,证明酶具有专一性。
- 关键提示:不宜选用碘液作为检测试剂,因为碘液只能检测淀粉是否被分解,无法检测蔗糖是否被水解。
影响酶活性的条件:该实验探究温度和 pH 对酶活性的影响,体现了酶作用条件较温和的特性。
- 探究温度对酶活性的影响:
- 材料选择:通常用淀粉酶。不能使用过氧化氢酶,因为 H_2O_2 本身受热会加速分解,干扰实验结果。
- 方法:设置 0℃、约 60℃ 和 100℃(沸水)三组温度。先将底物和酶分别在相应温度下保温后再混合。
- 检测:使用碘液。在最适温度下淀粉被完全水解,滴加碘液不变蓝;在高温或低温下淀粉未完全水解,滴加碘液变蓝。
- 探究 pH 对酶活性的影响:
- 材料选择:常用过氧化氢酶。
- 方法:设置酸性(HCl)、中性(蒸馏水)和碱性(NaOH)三个梯度的溶液环境。
- 现象:在中性(或最适 pH)环境下,产生气泡最多;在强酸或强碱环境下,基本无气泡产生。
- 核心结论:
- 酶在最适温度和最适 pH 下活性最高。
- 失活与抑制:高温、过酸、过碱会破坏酶的空间结构,导致酶永久失活;而低温会抑制酶的活性,但空间结构稳定,在适宜温度下活性可恢复。
这些实验加深了我们对酶作为生物催化剂本质的理解。
酶的基本概念
能量的释放、储存和利用,都必须通过化学反应来实现。细胞中每时每刻都进行着许多化学反应,统称为细胞代谢(cellular metabolism)。细胞代谢离不开酶(enzyme)。
加热能促进过氧化氢分解,是因为加热使过氧化氢分子得到了能量,从常态转变为容易分解的活跃状态。分子从常态转变为容易发生化学反应的活跃状态所需要的能量称为活化能(activation energy)。
Fe3+ 和过氧化氢酶能促进过氧化氢分解,它们没有给过氧化氢分子提供能量,而是降低了过氧化氢分解反应的活化能。如果把化学反应比作驾车翻越一座高山,“加热”相当于给汽车加大油门,用催化剂则相当于帮司机找到穿山的隧道。与无机催化剂相比,酶降低活化能的作用更显著,催化效率更高。正是由于酶的催化作用,细胞代谢才能在温和条件下快速有序地进行。
酶的化学本质不同于无机催化剂。一般来说,酶是活细胞产生的具有催化作用的有机物,其中绝大多数酶是蛋白质。酶的催化作用与无机催化剂有什么不同呢?
酶具有高效性。大量的实验数据表明,酶的催化效率是无机催化剂的 107 ~ 1013 倍。无机催化剂催化的化学反应范围比较广。例如,酸既能催化蛋白质水解,也能催化脂肪水解,还能催化淀粉水解。酶能像无机催化剂一样,催化多种化学反应吗?目前已发现的酶有 8 000 多种,它们分别催化不同的化学反应。
细胞中几乎所有的化学反应都是由酶催化的。酶催化特定化学反应的能力称为酶活性(enzyme activity)。酶活性可用在一定条件下酶所催化某一化学反应的速率表示。
实验过程中的变化因素称为变量。其中人为控制的对实验对象进行处理的因素叫作自变量,上述实验中加热、加 FeCl3 溶液、加肝脏研磨液,是对过氧化氢溶液的不同处理,温度和催化剂都属于自变量;因自变量改变而变化的变量叫作因变量,上述实验中过氧化氢分解速率就是因变量。除自变量外,实验过程中还存在一些对实验结果造成影响的可变因素,叫作无关变量。如上述实验中反应物的浓度和反应时间等。除作为自变量的因素外,其余因素(无关变量)都保持一致,并将结果进行比较的实验叫作对照实验。对照实验一般要设置对照组和实验组,上述实验中的 1 号试管就是对照组,2 号、3 号和 4 号试管是实验组。本实验的对照组未作任何处理,这样的对照组叫作空白对照。想一想,设置这样的对照组有什么意义?
过氧化氢酶只能催化过氧化氢分解,不能催化其他化学反应。脲酶除了催化尿素分解,对其他化学反应也不起作用。每一种酶只能催化一种或一类化学反应。细胞代谢能够有条不紊地进行,与酶的专一性是分不开的。许多无机催化剂能在高温、高压、强酸或强碱条件下催化化学反应。
溶液的温度和 pH 都对酶活性有影响。与无机催化剂相比,酶所催化的化学反应一般是在比较温和的条件下进行的。科学家采用定量分析的方法,分别在不同的温度和 pH 条件下测定同一种酶的活性,根据所得到的数据绘制成曲线图。分析这两个曲线图可以看出,在最适宜的温度和 pH 条件下,酶的活性最高。温度和 pH 偏高或偏低,酶活性都会明显降低。
一般来说,动物体内的酶最适温度在 35 ~ 40 ℃ ;植物体内的酶最适温度在 40 ~ 50 ℃ ;细菌和真菌体内的酶最适温度差别较大,有的酶最适温度可高达 70 ℃。动物体内的酶最适 pH 大多在 6.5 ~8.0,但也有例外,如胃蛋白酶的最适 pH 为 1.5;植物体内的酶最适 pH 大多为 4.5 ~ 6.5。
过酸、过碱或温度过高,会使酶的空间结构遭到破坏,使酶永久失活。在 0 ℃左右时,酶的活性很低,但酶的空间结构稳定,在适宜的温度下酶的活性会升高。因此,酶制剂适宜在低温下保存。
20 世纪 60 年代以前,医院里用的葡萄糖是用盐酸催化淀粉水解的方法来生产的,生产过程需要在 245 kPa 的高压和 140~150 ℃的高温下进行,并且需要耐酸的设备。60 年代以后改用酶法生产。
细胞中的各类化学反应之所以能有序进行,还与酶在细胞中的分布有关。拿植物叶肉细胞来说,与光合作用有关的酶分布在叶绿体内,与呼吸作用有关的酶分布在细胞质基质和线粒体内,这样,光合作用与呼吸作用在细胞内不同的区室同时进行,就可以互不干扰了。
酶的应用
这些课本中提到的酶的应用,展示了酶作为生物催化剂如何通过其高效性和专一性,在医疗、工业和环保等领域发挥巨大作用。以下是详细的展开讲解:
医疗卫生,抗菌、助消化与溶栓:
- 溶菌酶(Lysozyme):这种酶能特异性地水解细菌细胞壁中肽聚糖的化学键(NAG 与 NAM 之间的糖苷键),使细菌吸水破裂死亡。在临床上,它常与抗生素联用产生协同效应,增强杀菌力。
- 助消化药物:多酶片包含胃蛋白酶、胰淀粉酶、胰脂肪酶和胰蛋白酶等。它们能帮助将食物中的蛋白质、淀粉和脂肪分解为可吸收的小分子,用于治疗消化不良。
- 创伤处理与溶栓:胰蛋白酶能去除伤口坏死组织,促进愈合。针对心肌梗死等血栓疾病,医生会使用组织型纤溶酶原激活剂(t-PA)或尿激酶,它们能激活纤溶酶原转化为纤溶酶,从而溶解血块。
工业生产,提升效率与品质:
- 食品加工:
- 果胶酶:能分解果肉细胞壁中的果胶,显著提高果汁的出汁率并使果汁更清亮。
- 凝乳酶(Chymosin):在奶酪生产中,它能使牛奶中的酪蛋白凝集,最初从牛胃提取,现在已能通过转基因微生物大规模生产。
- 日化用品:加酶洗衣粉中添加了蛋白酶、脂肪酶和淀粉酶。为了使其在漂白剂存在下仍能保持活性,科学家利用蛋白质工程将枯草杆菌蛋白酶(Subtilisin)中易被氧化的氨基酸(如 222 位甲硫氨酸)替换,从而显著提升了酶的稳定性。
环境保护与绿色能源:
- 废物处理:脂肪酶能催化废弃油脂转化成生物柴油,实现资源合理利用并保护环境。
- 造纸与降解:在造纸工业中,利用酶替代化学法进行木质素降解和纸张漂白,不仅降低成本,还减少了环境污染。
酶工程对自然的改造。自然界中存在的酶有时在高温、强酸碱或有机溶剂下容易失活:
- 稳定性改造:例如利用定点突变技术在 T4 溶菌酶中引入二硫键,显著提高了其耐热性。
- 抗药性改造:青霉素酰化酶可以将天然青霉素改造成氨苄青霉素等杀菌力更强、广谱性更好的半合成抗生素。
酶的应用是否让你感受到了生物技术的奇妙?
酶抑制剂概述
酶抑制剂是指能够与酶分子结合并降低其催化活性的化学物质,它们在调节细胞代谢、药物开发及农业生产中具有核心地位。根据抑制剂与酶结合的性质及动力学特征,通常将其分为以下类别:
可逆抑制剂(Reversible Inhibitors):此类抑制剂通过非共价键与酶结合,抑制作用是暂时的,可以通过物理方法(如透析)移除。
竞争性抑制剂(Competitive Inhibitor):
- 机制:其分子结构通常与底物非常相似,因此能与底物竞争酶的活性位点。
- 特征:这种抑制是可逆的,增加底物浓度可以减弱甚至消除抑制作用。
- 动力学参数:最大反应速率(V_{max})保持不变,但米氏常数(K_m)增大(意味着酶与底物的表观亲和力下降)。
- 典型案例:磺胺类药物竞争性抑制细菌叶酸合成酶;丙二酸抑制琥珀酸脱氢酶。
非竞争性抑制剂(Non-competitive Inhibitor):
- 机制:抑制剂结合在酶活性位点之外的别构位点(Allosteric site),引起酶的空间结构改变。
- 特征:这种构象变化使得底物无法再与活性中心契合,或使已结合的 ES 复合物无法生成产物,因此无法通过增加底物浓度来解除。
- 动力学参数:V_{max} 减小,K_m 不变。
- 典型案例:重金属离子(如汞、铅)通过结合酶的侧链基团使酶失活。
反竞争性抑制剂(Uncompetitive Inhibitor):
- 机制:抑制剂仅与“酶 - 底物复合物”(ES)结合,而不结合游离酶。
- 动力学参数:V_{max} 和 K_m 同时减小。
不可逆抑制剂(Irreversible Inhibitors):这类抑制剂通常与酶活性中心的必需基团形成牢固的共价键,导致酶永久失活。
- 特征:抑制程度随时间增加而加强,无法通过物理方法除去,效果相当于降低了系统中有效酶的浓度。
- 典型案例:
- 有机磷农药/神经毒气:特异性结合乙酰胆碱酯酶的丝氨酸残基,导致神经系统信号传导中断。
- 青霉素:不可逆地抑制细菌合成细胞壁的关键酶,使细菌吸水破裂死亡。
在细胞代谢中,酶抑制剂不仅是外源药物,也是天然的调节工具。
- 机制:代谢途径的终产物(End product)往往作为抑制剂(通常是别构抑制),作用于该途径起始阶段的一个或多个酶。
- 意义:这是一种精密的负反馈调节(反馈抑制),能防止产物过量积累,确保细胞代谢有序进行。
许多现代药物本质上就是特定酶的抑制剂:
- 他汀类药物(Statins):通过抑制 HMG-CoA 还原酶来降低胆固醇水平。
- 格列卫(Gleevec):特异性结合超活化激酶的 ATP 结合区,阻断导致白血病的癌症信号。
这些抑制机制展示了生物分子之间精妙的契合与调节。
呼吸作用
新陈代谢
在深入探讨新陈代谢(Metabolism)这一生命最基本的特征时,我们需要从分子逻辑、细胞机制以及人体系统协调这三个层面进行重构。新陈代谢不仅是生物体内所有化学反应的总称,更是生命系统维持高度有序状态、抵御熵增(无序化)的核心手段。
在细胞水平上,新陈代谢由两套方向相反、但又精密耦合的化学反应组成:合成代谢(同化作用)与分解代谢(异化作用)。
- 分解代谢(Catabolism):这是一个“降解”的过程。细胞将复杂的有机大分子(如糖类、脂肪、蛋白质)分解为更简单的分子(如二氧化碳、水、尿素等)。
- 核心功能:释放储存在化学键中的能量,并将其捕获到活化载体分子(主要是 ATP)中;同时产生合成反应所需的“结构单元”(小分子)。
- 合成代谢(Anabolism):这是一个“构建”的过程。细胞利用分解代谢提供的能量和原料,合成自身所需的蛋白质、核酸、多糖等复杂大分子。
- 热力学本质:生命是一个开放的“耗散结构”。由于合成反应是熵减的过程(有序性增加),它必须与分解反应释放的能量偶联,并向环境排出热量(熵增),以符合热力学第二定律。
细胞内几乎所有的需能过程都不直接使用葡萄糖,而是通过 ATP(腺苷三磷酸) 这一能量货币。
- 结构特性:ATP 含有两个不稳定的高能磷酸键(磷酐键),末端磷酸基团具有较高的转移势能。
- 能量偶联:细胞通过 ATP 的水解(释放能量)来驱动吸能反应(如主动运输、肌肉收缩、物质合成);同时通过细胞呼吸(放能反应)将 ADP 和磷酸重新合成 ATP。
细胞代谢之所以能在常温、常压的温和环境下高效、有序进行,完全依赖于酶(Enzymes)。
- 作用机理:酶通过显著降低化学反应所需的活化能,使原本极其缓慢的反应在瞬间完成。
- 特性:具有极高的催化效率、专一性(一酶催一反应)以及对环境(pH、温度)的高度敏感性。
细胞呼吸是细胞获取能量的主要途径,主要分为有氧呼吸和无氧呼吸。
- 有氧呼吸(主要途径):在氧气参与下,葡萄糖被彻底氧化分解为 CO_2 和 H_2O。
- 场所:始于细胞质基质(糖酵解),完成于线粒体(柠檬酸循环与电子传递链)。
- 产能:1 分子葡萄糖彻底氧化可产生约 32 个 ATP。
- 无氧呼吸:在缺氧条件下,葡萄糖不完全分解产生乳酸或酒精,释放少量能量。人体在剧烈运动时,肌肉细胞会进行乳酸发酵以提供紧急能量。
人体的新陈代谢不是细胞活动的简单加和,而是由多个器官系统协同完成的物质与能量流转过程。物质摄取与废物排出的链条:
- 消化系统:将食物大分子分解为单糖、氨基酸、脂肪酸,这是代谢的第一阶段(消化阶段)。
- 呼吸系统:为细胞提供电子最终受体(O_2),并排出代谢废气(CO_2)。
- 循环系统:充当“物流网络”,将营养物质和氧气运送至全身组织细胞,同时将代谢废物运往排泄器官。
- 泌尿系统与皮肤:负责排出大部分含氮废物(如尿素)、多余的水和无机盐,维持内环境的水盐平衡。
代谢调节与内环境稳态(Homeostasis):人体内环境(血浆、组织液等)的理化性质(如血糖浓度、温度、pH)必须保持相对稳定,新陈代谢才能正常进行。
- 器官分工:
- 肝脏:代谢的中心实验室,调节血糖浓度,将有毒的氨转化为尿素。
- 肌肉:产热的主要来源(战栗产热与运动产热),高强度代谢消耗大量 ATP。
- 神经网络调节:
- 体液调节:二氧化碳浓度升高会刺激化学感受器,调节呼吸运动的深度和频率。
- 激素调节:胰岛素和胰高血糖素通过拮抗作用维持血糖稳态。甲状腺激素则能提高细胞代谢的速率,促进生长发育。
如果新陈代谢或其调节机制出现障碍,会导致一系列疾病。例如,肾功能衰竭(尿毒症)会导致代谢废物在内环境积聚,引起全身性中毒;糖尿病则是由于糖代谢调节受损导致的稳态失调。
新陈代谢是生命在微观分子逻辑上的高度精准耦合(由酶和 ATP 驱动)与在宏观系统层面的高度协调一致(由神经-体液-免疫调节维持)的统一体。它不仅是生物体生长、增殖的基础,更是生命之所以能成为“活”的系统的根本原因。
能量货币 ATP
三磷酸腺苷(ATP)被形象地称为细胞的“能量货币”,是生物体内最直接、最重要的能量来源。ATP 的化学本质是一种核苷酸,由三个部分组成:
- 腺嘌呤(Adenine):一个含氮碱基。
- 核糖(Ribose):一个五碳糖,与腺嘌呤结合形成腺苷(Adenosine)。
- 磷酸基团(Phosphate groups):三个磷酸基团依次连接在核糖上,分别称为 \alpha、\beta 和 \gamma 磷酸。
“高能”的秘密,磷酸酐键。ATP 分子中连接磷酸基团的化学键被称为磷酸酐键(常俗称为“高能磷酸键”,用“~”表示):
- 不稳定性:由于三个磷酸基团在生理 pH 下都带有负电荷,它们之间存在强烈的静电排斥力,这使得 ATP 分子极不稳定。
- 能量释放:当末端的磷酸键断裂(水解)时,会释放出大量的自由能。在活细胞内,1 mol ATP 水解为 ADP 和 Pi 释放的能量约为 57 kJ/mol,几乎是标准条件下(30.5 kJ/mol)的两倍。
ATP-ADP 循环,细胞的充电电池。ATP 在细胞内的含量非常低,但它通过与 ADP(腺苷二磷酸)之间的快速转化来满足巨大的能量需求:
- 水解(放能):ATP \rightarrow ADP + Pi + 能量。此过程为细胞的主动运输、肌肉收缩等提供动力。
- 再生(储能):ADP + Pi + 能量 \rightarrow ATP。能量主要来源于有机物的氧化分解(呼吸作用)或光合作用。
- 动态平衡:在一个典型的细胞中,ATP 分子每 1-2 分钟就会被更新一次,人体每天水解并再合成的 ATP 总量甚至相当于自身的体重。
能量利用的核心,能量偶联与磷酸化。细胞利用 ATP 的核心策略是能量偶联(Energy Coupling),即利用 ATP 水解这一放能反应来驱动原本无法自发进行的吸能反应:
- 磷酸化过程:ATP 水解时,末端磷酸基团往往不是简单漂走,而是转移到另一个分子上,这一过程称为磷酸化。
- 改变构象:受体分子(如离子泵蛋白)在磷酸化后处于高能、不稳定的状态,从而发生构象改变,完成特定的生物学功能(如 Na⁺/K⁺ 泵的转运)。
广泛的应用实例:
- 主动运输:维持细胞内外的离子浓度梯度。
- 机械功:驱动肌肉纤维收缩、纤毛摆动或分子马达在细胞骨架上行走。
- 生物合成:连接单体以构建蛋白质、核酸等生物大分子。
- 特殊功能:如萤火虫发光(化学能转化为光能)、电鳐放电以及大脑思考。
细胞中绝大多数需要能量的生命活动都是由 ATP 直接提供能量的,如大脑思考、电鳐发电和物质的主动运输都需要消耗 ATP。ATP 水解释放的能量是如何用于上述各种生命活动的呢?下图向你展示了 ATP 是如何为主动运输供能的。
参与 Ca2+ 主动运输的载体蛋白是一种能催化 ATP 水解的酶。当膜内侧的 Ca2+ 与其相应位点结合时,其酶活性就被激活了。
在载体蛋白这种酶的作用下,ATP 分子的末端磷酸基团脱离下来与载体蛋白结合,这一过程伴随着能量的转移,这就是载体蛋白的磷酸化。
载体蛋白磷酸化导致其空间结构发生变化,使 Ca2+ 的结合位点转向膜外侧,将 Ca2+ 释放到膜外。
ATP 水解释放的磷酸基团使蛋白质等分子磷酸化,这在细胞中是常见的。这些分子被磷酸化后,空间结构发生变化,活性也被改变,因而可以参与特定的化学反应。
细胞的能量梳理
细胞内的化学反应可以分成吸能反应和放能反应两大类。前者是需要吸收能量的,如蛋白质的合成等;后者是释放能量的,如葡萄糖的氧化分解等。许多吸能反应与 ATP 水解的反应相联系,由 ATP 水解提供能量;许多放能反应与 ATP 的合成相联系,释放的能量储存在 ATP 中,用来为吸能反应直接供能。也就是说,能量通过 ATP 分子在吸能反应和放能反应之间流通。因此,可以形象地把 ATP 比喻成细胞内流通的能量“货币”。正是由于细胞内具有 ATP 这种能量“货币”,才能及时而持续地满足细胞各项生命活动对能量的需求。
我们现在来梳理一下:
直接能源物质:ATP。
主要能源物质:葡萄糖。
生物体内良好的储能物质:脂肪。
动物细胞内的储能物质:糖原。
植物细胞内的储能物质:淀粉。
最终能量来源:太阳(光)能。
对比实验:设置两个或两个以上的实验组,通过对结果的比较分析,来探究某种因素对实验对象的影响,这样的实验叫作对比实验,也叫相互对照实验。在本节课的探究活动中,需要设置有氧和无氧两种条件,探究酵母菌在不同氧气条件下细胞呼吸的方式,这两个实验组的结果都是事先未知的,通过对比可以看出氧气条件对细胞呼吸的影响。对比实验也是科学探究中常用的方法之一。
细胞供能的核心在于 ATP(腺苷三磷酸),它被形象地称为细胞的“能量货币”。细胞通过将有机物(主要是葡萄糖)氧化分解,把释放的能量转移到 ATP 中,从而驱动各项生命活动。根据氧气的参与情况,细胞供能主要分为以下几种方式:
有氧呼吸(需氧呼吸)是绝大多数生物获取能量的主要途径,能将葡萄糖彻底氧化分解,释放大量能量。该过程分为三个阶段:
- 糖解途径(糖酵解):在细胞质基质中进行。1 分子葡萄糖分解为 2 分子丙酮酸,产生少量的 [H](还原型辅酶 I)和 2 分子 ATP(净产出)。
- 柠檬酸循环(三羧酸循环):在线粒体基质中进行。丙酮酸进入线粒体分解为 CO_2 和 [H],并产生少量 ATP。每分子乙酰辅酶 A 进入循环可产生 3 分子 NADH、1 分子 FADH_2 和 1 分子 GTP(等同于 ATP)。
- 氧化磷酸化:在线粒体内膜上进行。前两阶段产生的 [H](NADH 和 FADH_2)通过电子传递链与氧结合生成水,释放大量能量,驱动 ATP 合酶合成 大量 ATP。
- 能量转化效率:1 mol 葡萄糖彻底氧化可释放 2870 kJ 能量,其中约 34% 左右储存在 ATP 中,其余以热能形式散失。
无氧呼吸(发酵):在缺氧条件下,细胞通过酶的催化将有机物不彻底分解,释放少量能量。
- 过程:第一阶段与有氧呼吸的糖酵解完全相同。第二阶段丙酮酸在不同酶的催化下转化为乳酸或酒精(并产生 CO_2),目的是再生糖酵解所需的 NAD^+。
- 类型:
- 乳酸发酵:见于乳酸菌、动物骨骼肌、马铃薯块茎等。
- 酒精发酵:见于酵母菌和大多数植物器官(如苹果、水稻根)。
- 能量产出:1 mol 葡萄糖分解成乳酸仅产生 2 分子 ATP。
除了葡萄糖,细胞还可以代谢脂质和蛋白质:
- 脂肪:水解为甘油和脂肪酸。脂肪酸通过 \beta-氧化 产生乙酰辅酶 A 进入柠檬酸循环,产能量远高于糖类。
- 蛋白质:水解为氨基酸,脱去氨基后的碳链可进入有氧呼吸的不同环节进行氧化。
- 磷酸肌酸(肌肉特有):作为能量的“黄金储备”,能迅速将能量转移给 ADP 来补充 ATP。
| 能源物质 | 主要途径 | 场所 | 产能(每分子) |
|---|---|---|---|
| 葡萄糖 | 有氧呼吸 | 细胞质基质、线粒体 | 30-32 ATP |
| 葡萄糖 | 无氧呼吸 | 细胞质基质 | 2 ATP |
| 脂肪酸 | \beta-氧化 | 线粒体 | ~108 ATP (以软脂酸为例) |
这些机制共同构成了细胞的能量代谢网络。你是否想针对其中某一个具体过程(例如线粒体中的质子梯度是如何形成的)进行更深入的探讨?下面我们先综述,然后在下一节分别详细讲解。
总结有氧呼吸
对于绝大多数生物来说,有氧呼吸是细胞呼吸的主要形式,这一过程必须有氧的参与。有氧呼吸的主要场所是线粒体。线粒体具有内、外两层膜,内膜的某些部位向线粒体的内腔折叠形成嵴,嵴使内膜的表面积大大增加。嵴的周围充满了液态的基质。线粒体的内膜上和基质中含有许多种与有氧呼吸有关的酶。有氧呼吸最常利用的物质是葡萄糖,其化学反应式可以简写成:
\ce {C6H12O6 + 6H2O + 6O2 ->[酶] 6CO2 + 12H2O + \text {能量}}
有氧呼吸的全过程十分复杂,可以概括地分为三个阶段,每个阶段的化学反应都有相应的酶催化。
第一个阶段是,1 分子的葡萄糖分解成 2 分子的丙酮酸,产生少量的 [H],并且释放出少量的能量。这一阶段不需要氧的参与,是在细胞质基质中进行的。
第二个阶段是,丙酮酸和水彻底分解成二氧化碳和 [H],并释放出少量的能量。这一阶段不需要氧直接参与,是在线粒体基质中进行的。
第三个阶段是,上述两个阶段产生的 [H],经过一系列的化学反应,与氧结合形成水,同时释放出大量的能量。这一阶段需要氧的参与,是在线粒体内膜上进行的。
这里的 [H] 是一种十分简化的表示方式。这一产生 [H] 的过程主要是指氧化型辅酶 Ⅰ(NAD +)转化成还原型辅酶 Ⅰ(NADH)。
概括地说,有氧呼吸是指细胞在氧的参与下,通过多种酶的催化作用,把葡萄糖等有机物彻底氧化分解,产生二氧化碳和水,释放能量,生成大量 ATP 的过程。同有机物在生物体外的燃烧相比,有氧呼吸具有不同的特点:有氧呼吸过程温和;有机物中的能量经过一系列的化学反应逐步释放;这些能量有相当一部分储存在 ATP 中。
总结无氧呼吸
除酵母菌以外,还有许多种细菌(如乳酸菌)能够进行无氧呼吸。此外,马铃薯块茎、水稻根、苹果果实等植物器官的细胞以及动物骨骼肌的肌细胞等,除了能够进行有氧呼吸,在缺氧条件下也能进行无氧呼吸。一般地说,无氧呼吸最常利用的物质也是葡萄糖。
1 mol 葡萄糖在分解成乳酸以后,只释放出 196.65 kJ 的能量,其中只有 61.08 kJ 的能量储存在 ATP 中,近 69% 的能量都以热能的形式散失了。人体肌细胞无氧呼吸产生的乳酸,能在肝脏中再次转化为葡萄糖。
无氧呼吸的全过程,可以概括地分为两个阶段,这两个阶段需要不同酶的催化,但都是在细胞质基质中进行的。第一个阶段与有氧呼吸的第一个阶段完全相同。第二个阶段是,丙酮酸在酶(与催化有氧呼吸的酶不同)的催化作用下,分解成酒精和二氧化碳,或者转化成乳酸。无论是分解成酒精和二氧化碳或者是转化成乳酸,无氧呼吸都只在第一阶段释放出少量的能量,生成少量 ATP。葡萄糖分子中的大部分能量则存留在酒精或乳酸中。
无氧呼吸的化学反应式可以概括为以下两种:
\ce {C6H12O6 ->[酶] 2C3H6O3 (\text {乳酸}) + \text {少量能量}}
\ce {C6H12O6 ->[酶] 2C2H5OH (\text {酒精}) + 2CO2 + \text {少量能量}}
酵母菌、乳酸菌等微生物的无氧呼吸也叫作发酵。产生酒精的叫作酒精发酵,产生乳酸的叫作乳酸发酵。像这样,在没有氧气参与的情况下,葡萄糖等有机物经过不完全分解,释放少量能量的过程,就是无氧呼吸。
有氧呼吸和无氧呼吸都属于细胞呼吸。细胞呼吸是指有机物在细胞内经过一系列的氧化分解,生成二氧化碳或其他产物,释放能量并生成 ATP 的过程。所有生物的生存,都离不开细胞呼吸释放的能量。
细胞呼吸除了能为生物体提供能量,还是生物体代谢的枢纽。例如,在细胞呼吸过程中产生的中间产物,可转化为甘油、氨基酸等非糖物质;非糖物质代谢形成的某些产物与细胞呼吸中间产物相同,这些物质可以进一步形成葡萄糖。蛋白质、糖类和脂质的代谢,都可以通过细胞呼吸过程联系起来。
细胞呼吸的原理在生活和生产中得到了广泛的应用。生活中,馒头、面包、泡菜等许多传统食品的制作,现代发酵工业生产青霉素、味精等产品,都建立在对微生物细胞呼吸原理利用的基础上。在农业生产上,人们采取的很多措施也与调节呼吸作用的强度有关。例如,中耕松土、适时排水,就是通过改善氧气供应来促进作物根系的呼吸作用,以利于作物的生长;在储藏果实、蔬菜时,往往需要采取降低温度、降低氧气含量等措施减弱果蔬的呼吸作用,以减少有机物的消耗。
代谢途径
代谢途径(metabolic pathway),在生物化学中,是生物体将一种物质通过一系列酶的顺序作用(酶促反应)而转化成另一种代谢物的反应序列,包括合成或分解的过程。代谢途径由多个连续的代谢反应相互连接起来,进而完成物质的合成或分解,属于化学反应的一种。细胞内不同代谢途径组成了代谢网络。底物是否进入代谢途径,要视乎细胞的需要,即合成代谢物及分解代谢物浓度的独特组合(流量控制反应的动力)。代谢途径包括主要的代学反应(一般都是需要酶的)令生物保持它的内环境稳态。
代谢途径一般都有以下特征:它们包括多个步骤,就像瀑布一样。首个步骤多数不能倒逆。其他步骤并不一定不能倒逆。大部分情况下,这些步骤可以视乎细胞的实际需要而倒逆。糖酵解是极好的例子:当葡萄糖进入细胞后,它立即会被三磷酸腺苷(ATP)磷酸化,在首个不能倒逆的步骤成为葡萄糖 -6-磷酸。这是要避免葡萄糖离开细胞。在过多脂类或蛋白质时,糖酵解会倒逆进行糖异生,以产生葡萄糖六磷酸盐作为糖原或淀粉储存。它们都是有调节的,一般是以反馈方式,或是以循环形式再次进行反应(如三羧酸循环)。真核生物的合成代谢及分解代谢途径,大都局限于细胞中的特定区域(细胞区室),或是以使用不同的酶或辅助因子来分开。

呼吸作用(英语:cellular respiration),是生物体细胞把有机物氧化分解并转化能量的化学过程,该过程也称为释放作用。无论是否为自养,细胞内完成生命活动所需要的能量都是来自呼吸作用。在真核细胞中,线粒体是与呼吸作用密切相关的细胞器,呼吸作用的几个关键性步骤都在其中进行。呼吸作用是一种酶促氧化反应。虽名为氧化反应,不论有否氧气参与,都可称作呼吸作用(这是因为在化学上,有电子转移的反应过程,皆可称为氧化)。有氧气参与时的呼吸作用,称之为有氧呼吸;没氧气参与的反应,则称为无氧呼吸。
呼吸作用的目的,是透过释放食物里之能量,以制造三磷酸腺苷,即细胞最主要的直接能量供应者。呼吸作用与氢氧的燃烧之间最大分别是:呼吸作用透过一连串的反应步骤产生能量(一部分储存于 ATP 中),而燃烧则是将能量一次性的释放。在呼吸作用中,三大营养物质:碳水化合物、蛋白质和脂质的基本组成单位──葡萄糖、氨基酸和脂肪酸,被分解成更小的分子,并透过数个步骤,将能量转移到烟酰胺腺嘌呤二核苷酸(即 NADH)中。最后经过一系列的电子传递链,NADH 与氧气共同作用生成水,而原本贮存在其中的能量,则转移到 ATP 分子上,以供生命活动使用。
包扎伤口时,需要使用透气的消毒纱布或 OK 绷等敷料。以避免厌氧菌的繁殖,如破伤风芽孢杆菌。这种病菌只能进行无氧呼吸,皮肤破损较深或被锈钉扎伤病菌就容易大量繁殖。利用麦芽糖,葡萄糖,粮食和酵母菌以及发酵罐等,在控制通气的情况下可以生产各种酒;利用淀粉醋酸杆菌或谷氨酸棒状杆菌以及发酵罐控制通气的情况下,可以生产食醋或味精。花盆里的土壤板结后空气不足会影响根系生长,需要及时松土透气,有利于植物更好吸收氧气。稻田定期排水,避免水稻幼根因缺氧而变黑腐烂。提倡慢跑等有氧运动的原因之一是不致因剧烈运动而导致氧的不足,二是肌细胞无氧呼吸产生大量乳酸。乳酸的大量积累会使肌肉酸胀乏力。
糖酵解
糖酵解(Glycolysis)是生物体获取能量的基础代谢途径,具有极高的保守性,几乎存在于地球上所有的细胞中,证明了它起源于生命史的早期。
糖酵解发生在细胞的细胞质基质(胞质溶胶)中,无论是原核生物还是真核生物均是如此。该过程不需要氧气,属于厌氧代谢,是细胞在缺氧环境下唯一的能量来源(如成熟的哺乳动物红细胞)。
糖酵解的 10 步反应通常被分为两个阶段:
准备阶段(能量投入期):
- 关键激活:葡萄糖进入细胞后,首先被己糖激酶磷酸化为 6-磷酸葡萄糖。这一步不仅消耗了 1 分子 ATP,还将带负电荷的糖分子“锁”在了细胞内。
- 限速步骤:由磷酸果糖激酶 (PFK) 催化的第三步反应是整个途径最重要的调节开关,它消耗第二分子 ATP,将糖分子对称化,为后续的裂解做准备。
- 裂解:1 分子 6 碳糖最终被拆分为 2 分子相互转化的 3 碳糖(如 3-磷酸甘油醛)。
产出阶段(能量回收期):
- 氧化脱氢:3-磷酸甘油醛被氧化,脱下的电子和质子转移给 NAD^+,生成 NADH。
- 底物水平磷酸化:这是糖酵解生成 ATP 的直接方式。高能中间产物(如 1,3-二磷酸甘油酸和磷酸烯醇式丙酮酸)直接将磷酸基团转移给 ADP 生成 ATP。由于此前裂解成了两个 3 碳分子,此阶段的所有产出都是双倍的。
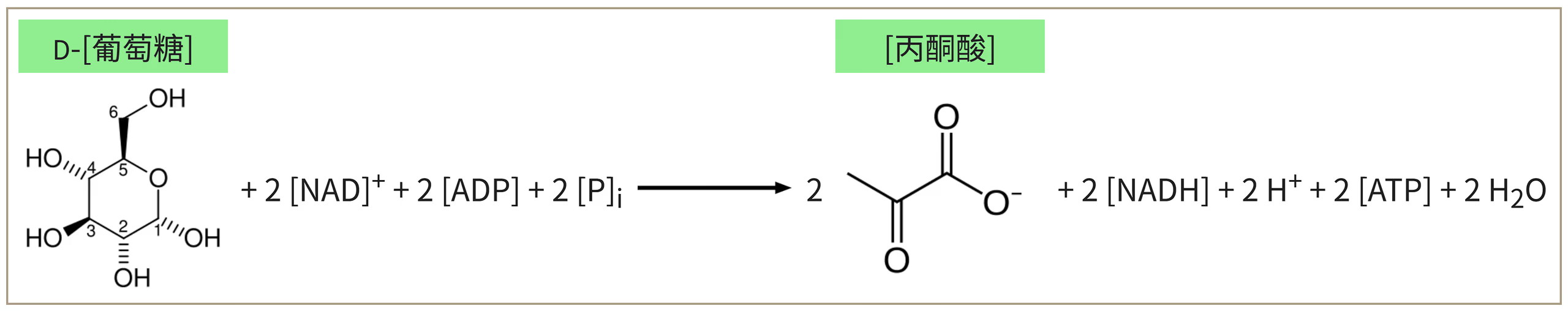
能量统计与代谢调节:
- 总账单:投入 2 ATP,产出 4 ATP 和 2 NADH。净收益为:2 ATP、2 NADH 和 2 丙酮酸。
- 精密调节:细胞根据能量状态实时调控。当 ATP 充足或柠檬酸积累时,会抑制 PFK 的活性,让糖酵解“减速”;当 ADP 或 AMP 增多时,则激活 PFK,加速产能。
糖酵解的终点是丙酮酸,它的命运由氧气决定:
- 有氧条件下:丙酮酸进入线粒体基质,氧化脱羧生成乙酰辅酶 A,进而参与三羧酸循环,产生大量的 ATP。
- 无氧条件下(发酵):为了让糖酵解不因为缺少 NAD^+ 而中断,细胞必须将 NADH 重新氧化。丙酮酸在不同酶的作用下转化为乳酸(动物、乳酸菌)或乙醇和 CO_2(酵母、多数植物)。
糖酵解不仅是产能过程,它产生的许多中间产物还为合成氨基酸、脂质和核苷酸提供了重要的碳骨架。
三羧酸循环
三羧酸循环(TCA 循环)是有氧呼吸的核心,它不仅是能量产生的“主发动机”,更是细胞内各种物质代谢的枢纽。以下是这一精密过程的详细解析:
循环的发生场所与“入场券”:在真核细胞中,这一循环完全发生在线粒体基质中。在循环开始前,糖酵解产生的丙酮酸必须通过线粒体膜进入基质,由丙酮酸脱氢酶复合体催化,脱去一个 CO_2 并产生一分子 NADH,最终转化为乙酰辅酶 A(Acetyl-CoA)。乙酰辅酶 A 就像一张“入场券”,将碳骨架送入循环。
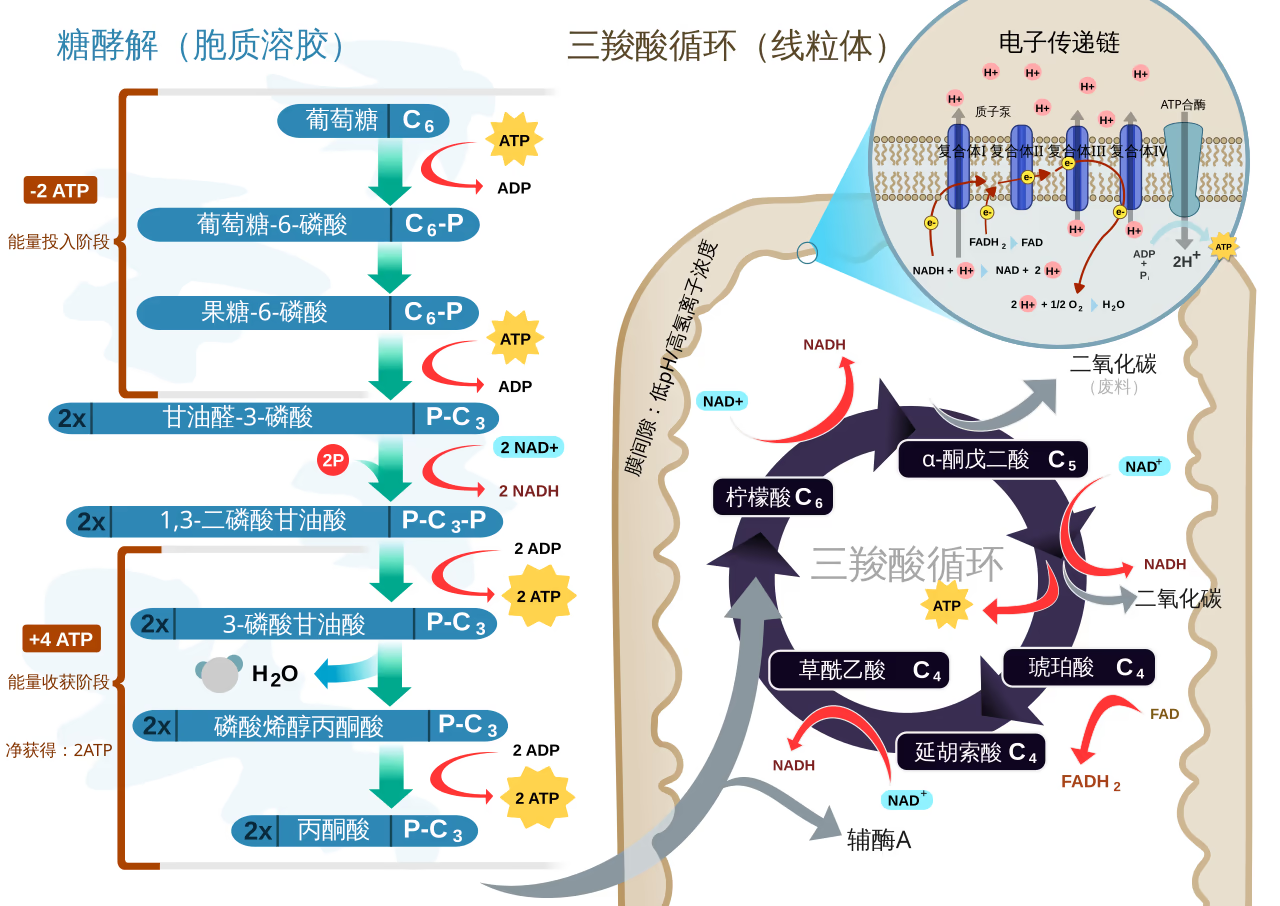
整个循环由 8 个特定的酶促反应组成,通过不断的脱氢和脱羧来提取能量:
- 起始缩合(第 1 步):乙酰辅酶 A (2C) 与草酰乙酸 (4C) 缩合成柠檬酸 (6C)。这是整个循环的开端,也是重要的调控点。
- 氧化脱羧(第 3-4 步):异柠檬酸经脱氢脱羧生成 \alpha-酮戊二酸,随后 \alpha-酮戊二酸再次氧化脱羧生成琥珀酰辅酶 A。这两步共释放了 2 分子 CO_2 并产生了 2 分子 NADH。
- 底物水平磷酸化(第 5 步):琥珀酰辅酶 A 转化为琥珀酸,直接生成一分子 GTP 或 ATP。这是循环中唯一直接产生高能磷酸键的步骤。
- 受氧限制的再生(第 6-8 步):琥珀酸经过脱氢(生成 FADH_2)、加水和再次脱氢(生成 NADH),最终重新生成草酰乙酸。虽然循环本身不消耗氧,但 NADH 和 FADH_2 的再生高度依赖于后续电子传递链对氧的利用。
能量统计与代谢产出:每消耗 1 分子乙酰辅酶 A(对应半个葡萄糖),循环的净产出为
- 3 分子 NADH 和 1 分子 FADH_2:携带高能电子进入电子传递链。
- 1 分子 ATP 或 GTP:直接可用的能量。
- 2 分子 CO_2:代谢废弃物。
调节机制与双向代谢枢纽:
- 精密调节:循环的速度受细胞能量状态(ATP/ADP 比率)调节。当 ATP 充足时,异柠檬酸脱氢酶等会被抑制,减缓生产;反之则被激活。
- 双向代谢(Amphibolic):TCA 循环不仅是分解途径(Catabolic),产生的中间产物如 \alpha-酮戊二酸和草酰乙酸常被抽走,用于合成氨基酸、脂质和核苷酸等(合成代谢/Anabolic)。
- 添补反应:为了防止中间产物被抽走导致循环停滞,细胞通过添补反应(如丙酮酸羧化为草酰乙酸)来及时补充“燃料”。
这些产生的还原辅酶(NADH 和 FADH_2)随后将进入氧化磷酸化阶段。
丙酮酸脱羧
丙酮酸脱羧是连接糖酵解与三羧酸循环的关键“桥梁反应”,在代谢调节中占据核心地位。
发生场所与核心装置:在真核细胞中,这一过程发生在线粒体基质中。催化这一反应的是一个巨大的丙酮酸脱氢酶复合体 (PDC),它由三种酶(E1、E2、E3)组成,其体积甚至比核糖体还要大。该复合体在氧化脱羧过程中需要多种维生素衍生物作为辅酶,包括来自维生素 B_1 的 TPP 和来自维生素 B_5 的辅酶 A (CoA)。
氧化脱羧的“三步走”过程:在 PDC 的精密配合下,1 分子丙酮酸(3C)最终转化为 1 分子乙酰辅酶 A(2C)。
- 脱羧与结合:丙酮酸首先脱去一个 CO_2,剩下的二碳部分与 TPP 结合形成羟乙基。
- 氧化脱氢:羟乙基被氧化为乙酰基,脱下的电子通过还原 NAD^+ 产生 NADH,为后续的氧化磷酸化储备高能电子。
- 生成乙酰辅酶 A:活化的乙酰基被转移到辅酶 A (CoA) 上,形成进入三羧酸循环的“入场券”——乙酰辅酶 A。
这一反应是细胞代谢的一个决定性时刻:
- 不可逆性:在动物体内,丙酮酸一旦转化为乙酰辅酶 A,就无法再通过糖异生变回葡萄糖。
- 枢纽作用:它不仅处理糖类代谢产物,脂肪酸的 \beta-氧化也会产生乙酰辅酶 A 进入此路径,使三羧酸循环成为三大营养物质氧化的共同通路。
- 去向选择:当细胞能量(ATP)短缺时,乙酰辅酶 A 进入循环彻底氧化;当能量充足时,它可转变为柠檬酸并回到细胞质中用于合成脂肪。
另一种路径:简单脱羧。在缺氧环境下的酒精发酵中,丙酮酸不进行氧化脱羧,而是由丙酮酸脱羧酶催化直接释放 CO_2 生成乙醛,随后被还原为乙醇。
这个反应结束后,乙酰辅酶 A 就要正式踏入三羧酸循环的大门了。对于这一步中涉及的多种辅酶(如维生素 B_1 或 B_5)的生化作用,或者后续的能量产出。
氧化磷酸化
氧化磷酸化(Oxidative Phosphorylation)是有氧呼吸的最终阶段,也是真核细胞产生 ATP 的主要途径。
这一精密过程可以拆解为两个阶段:
电子传递链(ETC)由一系列嵌入线粒体内膜的蛋白质复合体组成。
- 能量来源:来自糖酵解和三羧酸循环的 NADH 和 FADH_2 贡献出高能电子。
- 质子泵送:当电子在复合体(I-IV)间逐级传递时,释放的能量将质子(H^+)从线粒体基质泵入膜间隙。
- 最终受体:电子最终传递给氧气(O_2),并结合质子生成水(H_2O)。
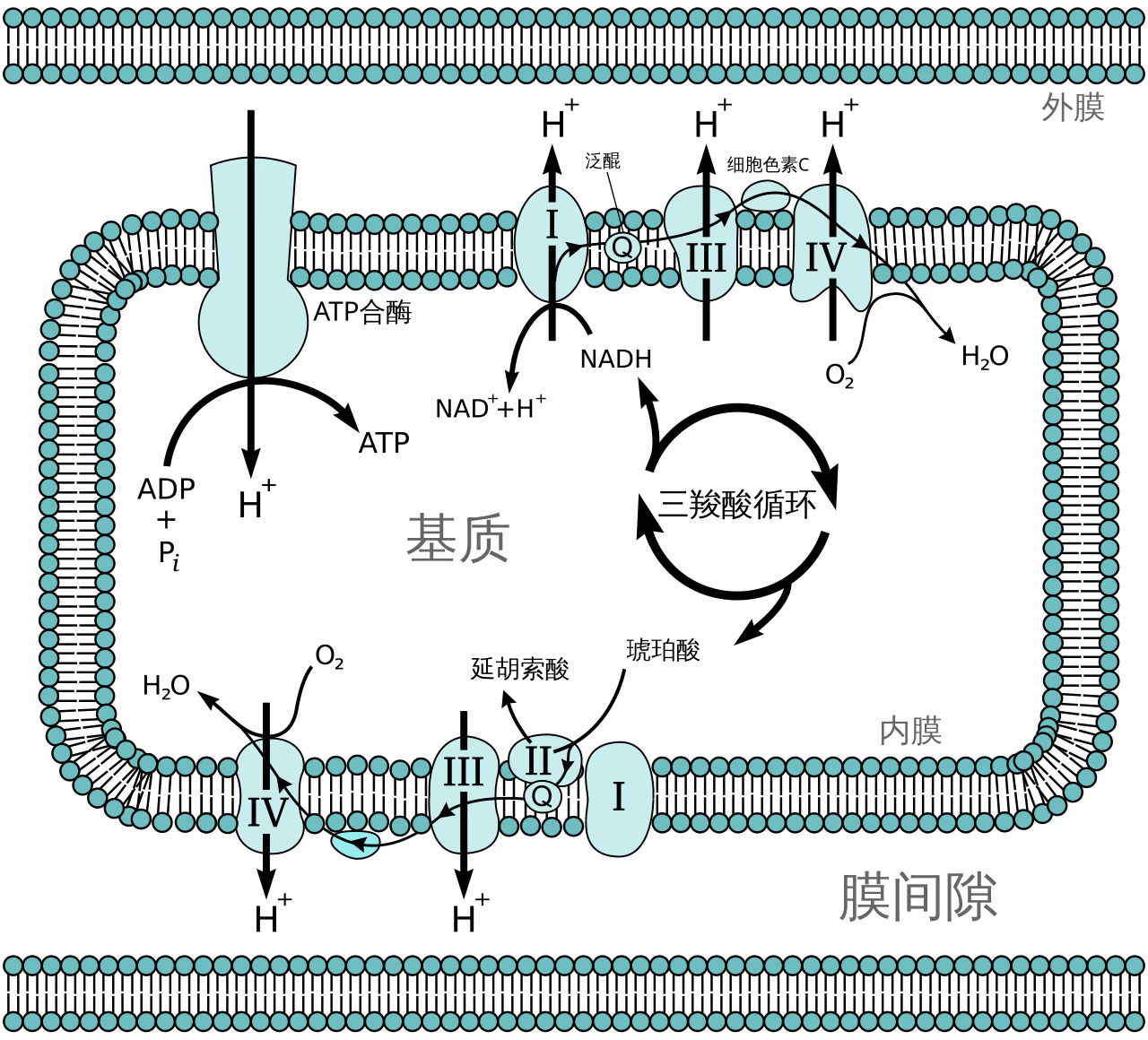
化学渗透(Chemiosmosis):质子在膜两侧形成了浓度梯度和电位差,这被称为质子动力势。
- ATP 合酶:质子顺着电化学梯度通过 ATP 合酶回流到基质中。
- 能量转化:ATP 合酶如同“分子涡轮”,利用质子流的能量将 ADP 和 Pi 合成为 ATP。
能量统计与调节:
- 高效产出:1 分子葡萄糖通过完整氧化分解,最终可产生约 30 或 32 个 ATP 分子。
- 抑制与调节:氰化物通过抑制细胞色素 c 氧化酶阻断电子传递;而解偶联蛋白(如在棕色脂肪中)能使质子不经 ATP 合酶流回,从而将能量直接转化为热能。
通过这一过程,细胞将营养物质中的化学能高效地转化为通用的“能量货币”。至此,我们已经完整梳理了从糖酵解到氧化磷酸化的呼吸全过程。
无氧发酵
无氧发酵(通常指无氧呼吸)是细胞在无氧条件下,通过酶的催化作用将有机物不彻底氧化分解,释放少量能量并产生 ATP 的过程。
这一过程完全发生在细胞质基质中,可以分为两个核心阶段:
无论是酒精发酵还是乳酸发酵,第一阶段完全相同:
- 物质变化:1 分子葡萄糖被分解为 2 分子丙酮酸,并产生少量的 [H](即 NADH)。
- 能量统计:此阶段释放少量能量,净产生 2 个 ATP。
由于细胞所含酶的种类不同,丙酮酸在第二阶段会有不同的去向:
酒精发酵 (Alcoholic Fermentation):
- 过程:丙酮酸首先在丙酮酸脱羧酶的作用下脱去 CO_2 生成乙醛,乙醛再被 NADH 还原为乙醇。
- 反应式:C_6H_{12}O_6 \xrightarrow{酶} 2C_2H_5OH + 2CO_2 + 少量能量。
- 典型生物:酵母菌以及大多数植物在缺氧时(如淹水的根系或苹果果实)。
乳酸发酵 (Lactic Acid Fermentation):
- 过程:丙酮酸在乳酸脱氢酶的催化下,直接被 NADH 还原为乳酸,此过程不产生 CO_2。
- 反应式:C_6H_{12}O_6 \xrightarrow{酶} 2C_3H_6O_3 + 少量能量。
- 典型生物:乳酸菌、高等动物剧烈运动时的骨骼肌,以及少数植物器官(如马铃薯块茎、玉米胚、甜菜块根)。
生理意义与核心机制:
- NAD^+ 再生:发酵的关键不在于产生更多能量,而在于将糖酵解产生的 NADH 重新氧化为 NAD^+,从而保证糖酵解能够持续进行并提供最基本的 ATP。
- 能量利用率低:葡萄糖中的大部分能量仍储存在酒精或乳酸中,未被完全释放。
β-氧化
\beta-氧化是脂肪酸分解的关键途径,它通过一种“螺旋式”的重复过程,将长链脂肪酸彻底拆解为可供能的单元。
在进入线粒体进行氧化之前,脂肪酸必须先在细胞质中完成活化。
- 活化与耗能:脂肪酸在脂肪酰辅酶 A 合成酶的作用下,与辅酶 A (CoA) 结合生成脂酰辅酶 A。这一过程需要消耗 ATP。
- 进入线粒体:长链脂酰辅酶 A 不能直接穿过线粒体内膜。它必须借助肉碱作为转运载体,通过内膜上的转运蛋白进入线粒体基质。
一旦进入基质,脂酰辅酶 A 就会经历由四种酶催化的循环反应。每经过一轮循环,碳链就会在 \beta 位被切断,缩短 2 个碳原子:
- 脱氢(氧化):脱下 2 个氢原子产生 FADH_2。
- 水合:引入一分子水,为下一步氧化做准备。
- 再脱氢(氧化):再次脱氢产生 NADH。
- 硫解:在硫解酶的作用下,切下一个 2 碳的乙酰辅酶 A。剩下的脂酰辅酶 A 则比原来少 2 个碳,并再次进入循环。
结果:高效的能量释放与碳架特征
- 能量产出:脂肪是细胞内最高效的“蓄电池”。1g 脂肪彻底氧化分解产生的能量(约 39 kJ)是等质量糖类(约 17 kJ)的 2 倍多。这是因为脂肪分子中氢含量更高,而氧含量远低于糖类。
- 碳数规律:由于该过程每次精准地剪下 2 个碳原子,这解释了为什么自然界中绝大多数生物合成的脂肪酸都含有偶数个碳原子。
产物的去向:生成的乙酰辅酶 A 随后会直接进入三羧酸循环(TCA 循环)彻底氧化。而循环中产生的 NADH 和 FADH_2 则携带高能电子进入电子传递链,通过氧化磷酸化产生大量的 ATP。
这一过程展示了细胞如何将“沉重”的脂肪转化为灵活的通用能量。
光合作用
在植物工厂里,人工光源可以为植物的生长源源不断地提供能量。在自然界,则是万物生长靠太阳。太阳光能的输入、捕获和转化,是生物圈得以维持运转的基础。光合作用(photosynthesis)是唯一能够捕获和转化光能的生物学途径。因此,有人称光合作用是“地球上最重要的化学反应”。无论是在植物工厂里,还是在自然界,植物捕获光能要依靠特定的物质和结构。
光合作用的强度(简单地说,就是指植物在单位时间内通过光合作用制造糖类的数量),直接关系农作物的产量,研究影响光合作用强度的环境因素很有现实意义。
根据光合作用的反应式可以知道,光合作用的原料 —— 水、CO2,动力 —— 光能,都是影响光合作用强度的因素。因此,只要影响到原料、能量的供应,都可能是影响光合作用强度的因素。例如,环境中 CO2 浓度,叶片气孔开闭情况,都会因影响 CO2 的供应量而影响光合作用的进行。叶绿体是光合作用的场所,影响叶绿体的形成和结构的因素,如无机营养、病虫害,也会影响光合作用强度。此外,光合作用需要众多的酶参与,因此影响酶活性的因素(如温度),也是影响因子。
绿叶中色素的提取和分离
对于高等植物来说,叶片是进行光合作用的主要器官。这些植物的叶片多数是绿色的,说明其中有绿色的色素。在玉米地里,有时可以看到叶片中不含绿色色素的白化苗。这样的白化苗,待种子中储存的养分耗尽就会死去。可见,叶片中的色素可能与光能的捕获有关。
课本实验“绿叶中色素的提取和分离”是高中生物中非常经典且重要的实验,它通过物理化学方法揭示了植物光合作用的物质基础。下面我为你详细解析实验的原理、步骤要点以及结果分析。实验原理:
- 提取原理:叶绿体中的色素属于有机物,不溶于水,但能溶解在有机溶剂中(如无水乙醇、丙酮等)。因此,我们利用无水乙醇作为提取液,将色素从破碎的细胞中“泡”出来。
- 分离原理:这涉及到纸层析法。不同色素在层析液(一种脂溶性很强的溶剂)中的溶解度不同。溶解度高的色素随层析液在滤纸上扩散得快;溶解度低的则扩散得慢。经过一段时间,原本混合在一起的色素就会因“跑”的速度不同而分离开来。
关键试剂的作用:
- 无水乙醇:作为提取剂,溶解色素。
- 二氧化硅(SiO₂):增加摩擦力,使研磨更加充分,从而破坏细胞结构释放色素。
- 碳酸钙(CaCO₃):极其重要。研磨时细胞中的有机酸会释放,酸性环境会破坏叶绿素分子(使其脱镁)。加入碳酸钙可以中和有机酸,保护色素不被破坏。
- 层析液:作为流动相,利用溶解度差异分离色素。
操作步骤的深度细节:
- 提取色素(研磨与过滤):
- 研磨要迅速:目的是防止乙醇挥发,并减少色素被空气氧化。
- 尼龙布过滤:不能用滤纸过滤,因为滤纸会大量吸附色素,导致滤液浓度过低。
- 制备滤纸条:
- 剪去两角:目的是防止层析液在滤纸边缘扩散过快,导致色素带呈现弧形而非整齐的直线。
- 画滤液细线:
- 要求:细、齐、直。
- 重复:待滤液干后,需重复画 2-3 次,目的是积聚更多的色素,使分离后的色素带更加清晰明显。
- 色素分离(层析):
- 关键点:层析液绝不能触及滤液细线。如果触碰,细线上的色素会直接溶解到烧杯里的层析液中,导致实验失败,滤纸条上什么也跑不出来。
- 密封:层析时需加盖,防止层析液中的有毒挥发性物质扩散到空气中。
实验结果:四条彩虹带。层析结束后,滤纸条上从上到下(即从扩散最快到最慢)依次出现四条色素带。
- 胡萝卜素:橙黄色,最快(溶解度最高),最窄(含量最少)。
- 叶黄素:黄色,次之。
- 叶绿素 a:蓝绿色,较慢,最宽(含量最多,约占 3/4 中的大部分)。
- 叶绿素 b:黄绿色,最慢(溶解度最低),较宽。
总结:这个实验成功与否,很大程度上取决于研磨时色素是否得到了充分保护(CaCO₃),以及画线和层析时操作的规范性。
叶绿素
叶绿素是光合作用中最核心的色素,主要存在于叶绿体的类囊体薄膜上。它不仅是植物呈现绿色的原因,更是将太阳能转化为化学能的关键“机器”。种类与分布:
- 主要类型:高等植物中主要含有叶绿素 a(蓝绿色)和叶绿素 b(黄绿色)。在所有光合放氧生物中,叶绿素 a 是必需的。
- 比例:在正常绿叶中,叶绿素的含量约占色素总量的 3/4,其中叶绿素 a 与叶绿素 b 的比例约为 3:1。
光谱吸收特性:
- 吸收区域:叶绿素主要吸收红光和蓝紫光。
- 叶绿素 a:吸收峰分别在波长 430 nm(蓝紫)和 662 nm(红)附近。
- 叶绿素 b:吸收峰分别在波长 453 nm(蓝)和 642 nm(红橙)附近。
- 为什么是绿色的:叶绿素对绿光的吸收最少,绿光会被反射或透过,因此我们看到的叶片呈现绿色。
分子结构与成分:
- 核心结构:叶绿素分子包含一个卟啉环,其中心含有一个镁原子(Mg)。
- 营养需求:镁是合成叶绿素的必需元素,若植物缺镁,叶绿素无法合成,叶片会发黄(失绿症)。
- 疏水尾部:分子末端有一条长长的疏水“尾巴”(叶醇基),使叶绿素分子能牢牢地锚定在类囊体薄膜的脂双层中。
生物学功能:
- 能量捕获(天线色素):绝大多数叶绿素分子像天线一样吸收并传递光能。
- 能量转换(反应中心):极少数特殊状态的叶绿素 a(如 P680 和 P700)构成反应中心。它们受光激发后会释放高能电子,实现光能向电能的转换,这是光反应的起始点。
环境影响因素:
- 光的诱导:叶绿素的形成需要光。避光生长的豆芽呈黄色(黄化现象),见光后才能合成叶绿素变绿。
- 季节变化:秋季气温降低,叶绿素分子变得不稳定并易被破坏,而类胡萝卜素较稳定,导致叶片显现出黄色。
叶绿素在光合作用中通过捕获光能,驱动了水分子的裂解和 ATP、NADPH 的合成,为后续合成糖类提供了动力。
叶绿体
叶绿体是绿色植物进行光合作用的核心细胞器,被誉为植物细胞的“养料制造车间”和“能量转换站”。
叶绿体通常呈扁平的椭球形或球形,由双层膜包被。其内部结构设计极大地增加了光合作用的有效面积:
- 类囊体与基粒:叶绿体基质中悬浮着许多扁平的囊状结构,称为类囊体。类囊体叠集成基粒,这种结构使光吸收面积最大化(1 克菠菜叶片的类囊体总面积可达约 60 平方米)。
- 基质:内膜与类囊体之间的浓稠液体,含有固定二氧化碳所需的酶,以及少量的 DNA、RNA 和核糖体。
光合色素:光的“捕获者”光合色素分布在类囊体薄膜上。主要分为两类:
- 叶绿素(约占 3/4):包括叶绿素 a(蓝绿色)和叶绿素 b(黄绿色),主要吸收蓝紫光和红光。
- 类胡萝卜素(约占 1/4):包括胡萝卜素(橙黄色)和叶黄素(黄色),主要吸收蓝紫光。
- 绿光反射:由于这些色素基本不吸收绿光,绿光被反射或透过,使叶片呈现绿色。
功能:能量转换的两个阶段。叶绿体通过两个相互联系的阶段将光能转化为化学能:
- 光反应(在类囊体薄膜进行):色素捕获光能,将水分解并释放氧气,同时产生 ATP 和 NADPH(活跃化学能)。
- 碳反应/暗反应(在基质中进行):利用光反应提供的 ATP 和 NADPH,将二氧化碳固定并还原为糖类(稳定的化学能)。
生物学特性:半自主与起源。叶绿体具有一定的遗传独立性,能合成部分自身所需的蛋白质。科学界普遍认为叶绿体起源于被古代真核细胞吞噬的光合细菌(内共生起源学说)。
实验验证:恩格尔曼通过水绵实验证明,叶绿体受光部位会释放氧气,直接证实了它是光合作用的场所。
光合作用概述
光合作用是生物界乃至整个自然界最基础的代谢过程。光合作用是指绿色植物、藻类以及某些细菌(如蓝细菌)利用光能,通过叶绿体或光合片层,将二氧化碳和水(或硫化氢等无机物)转化成储存着能量的有机物(如糖类),并释放出氧气(或硫)的过程。根据产物不同,可分为产氧光合作用(利用水并释放氧气)和不产氧光合作用(如某些细菌利用硫化氢并释放硫)。
人类对光合作用的认识经历了从直观推测到定量实验,再到分子水平研究的漫长过程:
- 土壤源泉说:亚里士多德曾认为植物生长所需的物质全部来自土壤。
- 柳树实验(1642 年):比利时人海尔蒙特通过五年的盆栽柳树实验发现,土壤质量几乎未减,从而推论植物生长主要来源于水而非土壤。
- 空气净化实验(1771 年):普里斯特利通过薄荷枝条和蜡烛、小鼠的实验,证明植物能“净化”被污浊的空气。
- 光的重要性(1779 年):英格豪斯发现只有植物的绿色部分在光照下才能净化空气。
- 能量转化(1845 年):迈尔根据能量守恒定律指出,植物在光合作用时把光能转换成化学能储存。
- 淀粉生成(1864 年):萨克斯通过叶片一半曝光、一半遮光的对比实验,证实光合作用产生了淀粉。
- 场所与光谱(1880 年):恩格尔曼利用水绵和好氧细菌证明,叶绿体是光合作用的场所,且红光和蓝紫光效率最高。
- 氧气来源(1941 年):鲁宾和卡门利用 ^{18}\text{O} 同位素标记法,确证光合作用释放的氧气全部来自水。
- 卡尔文循环(1945-1957 年):卡尔文用 ^{14}\text{C} 标记二氧化碳,探明了碳在光合作用中转化成有机物的具体途径。
光合作用被誉为“地球上最重要的化学反应”,其意义远超生物学范畴:
- 物质与能量源泉:它为生物圈中几乎所有生物提供食物和能量来源。
- 维持碳氧平衡:通过吸收二氧化碳、释放氧气,光合作用对维持大气成分的相对稳定和减缓温室效应起着至关重要的作用。
- 化石燃料的根基:人类目前使用的煤炭、石油、天然气,本质上都是古代植物通过光合作用固定的太阳能。
- 演化推动力:由于蓝细菌等早期光合生物释放氧气,才催生了需氧生物的繁衍和陆生生物的出现。
化能合成作用
化能合成作用(Chemosynthesis)是自养生物(主要是原核生物)不利用光能,而是利用体外环境中的无机物氧化时所释放的化学能,将二氧化碳和水合成糖类的过程。
与光合作用类似,化能合成作用也是一个氧化还原过程,但其能量来源完全不同:
- 能量捕获:生物通过氧化还原电位较高的无机物(如 NH_3、H_2S、Fe^{2+}、H_2)获取高能电子。
- 碳固定:利用氧化释放的化学能推动类似卡尔文循环的反应,将 CO_2 还原为有机物(如糖类),并储存能量。
- 生态地位:这些生物是稀有生态系统(如深海、黑洞穴)中的生产者,支撑着不依赖阳光的生物圈。
模式生物及其生化反应:
- 硝化细菌:包括亚硝酸细菌(如Nitrosomonas)和硝酸细菌(如Nitrobacter)。它们在氮循环中至关重要,能将植物难以利用的铵盐转化为易吸收的硝酸盐。
- 硫细菌:例如Beggiatoa,能在黑暗中氧化 H_2S 产生单质硫。在深海热泉,这类细菌是管虫、贻贝等生物的共生伙伴,为其提供营养。
- 产甲烷古菌:这是一类极其古老的化能自养生物。例如Methanococcus jannaschii,它利用从热水口排放的 H_2 还原 CO_2 产生甲烷,并在此过程中泵送质子以合成 ATP。
- 氢细菌:利用分子氢氧化产生的能量同化二氧化碳,通常为兼性化能自养菌。
“生命摇篮”深海热泉与进化?化能合成作用被认为是地球上最古老的生物自养过程,甚至早于光合作用:
- 代谢起源说:该假说认为生命起源于热液喷口附近的化学反应,逐渐发展出基于质子梯度的代谢结构。
- 极端环境适应:这些生物多为嗜极生物(Extremophiles),能在高温(可达 350℃)、高压、强酸或高盐的极端环境下生存,反映了早期地球的生存条件。
- 大气重塑的前奏:在产氧光合生物出现前,化能自养生物是地球化学循环的主导者,它们将碳元素从大气转移至岩石圈(形成碳酸盐),对调节地球早期气温起到了作用。
化能合成细菌在现代生物技术中也有重要应用:
- 环境修复:利用嗜油细菌降解原油污染,或利用工程菌吸收环境中的重金属。
- 湿法冶金:利用微生物的氧化还原特性,将矿石中的金属溶解并提取出来,具有低污染、高回收的优点。
化能合成作用展示了生命极强的适应性。
探究环境因素对光合作用强度的影响
这份探究实验的核心是通过小圆形叶片浮起法(叶片真空渗入法)来定量测量光合作用强度。其背后的生物学逻辑非常严密,我们可以从实验设计、原理和环境因子三个维度来深度拆解:
- 抽气处理(关键步骤):利用注射器造成负压,将叶肉细胞间隙中的空气排出,代之以液体,使叶片由于密度增大而沉入水底。
- CO_2 的定额供应:实验中使用 NaHCO_3 溶液(通常为 1%~2%)。它在水中充当 CO_2 缓冲液,能持续稳定地提供光合作用所需的原料,保证 CO_2 浓度不成为限制实验的变量。
- 自变量的控制:通过改变台灯与烧杯的距离来调节光照强度。距离越近,单位面积接收的光子越多,光反应产生的 [H] 和 ATP 就越多。
为什么“叶片浮起”能代表光合速率?
- 浮起机制:叶片沉在水底时只进行呼吸作用。一旦光照开启,光反应裂解水产生的 O_2 会填充进叶肉细胞的间隙。随着 O_2 的积累,叶片浮力增大,最终浮起。
- 净光合速率:注意,实验测得的是净光合速率。因为在叶片制造 O_2 的同时,线粒体也在消耗 O_2 进行呼吸作用。只有当光合产生的 O_2 超过呼吸消耗的 O_2 时,多余的气体才会让叶片浮起。因此,单位时间内浮起的叶片数越多,说明净光合强度越大。
影响光合强度的“三驾马车”除了光照强度,你还可以根据这个实验框架更换变量:
- CO_2 浓度:配置不同浓度的 NaHCO_3 溶液。在一定范围内,光合速率随 CO_2 浓度升高而增加,直到达到饱和。
- 温度:使用恒温水浴。温度主要通过影响酶的活性来影响暗反应(卡尔文循环)的速率。
- 光质(光色):在台灯上罩不同颜色的玻璃纸。由于叶绿素主要吸收红光和蓝紫光,在这两种光下叶片浮起的速度通常比绿光下快得多。
实验异常现象分析:
- 光饱和点:如果距离缩短到一定程度后,叶片浮起速度不再加快,说明已达到光饱和点。此时限制因素可能是 CO_2 浓度或温度。
- 光补偿点:如果光线极弱,叶片可能永远不会浮起。这意味着此时的光合强度小于或等于呼吸强度,即处于或低于光补偿点。
光反应阶段
光反应概述
一般来说,光合作用是指绿色植物通过叶绿体,利用光能,将二氧化碳和水转化成储存着能量的有机物,并且释放出氧气的过程。这一过程可以用下面的化学反应式来概括,其中(CH₂O)表示糖类。
\ce {CO2 + H2O ->[光能][叶绿体] (CH2O) + O2}
自然界中到处都有二氧化碳、水和阳光,然而,能够利用它们合成有机物的却只有进行光合作用的细胞。完成这一神奇过程的就是叶绿体。
在大学生物学中,高中所学的“光反应”和“暗反应”会被拆解为更精确的分子机制和代谢途径。光反应在大学阶段通常被称为 光合电子传递反应,它主要对应两种电子流动方式:
- 非循环电子传递(Non-cyclic electron flow):这是光反应的主要形式,涉及光系统 II(PSII)和光系统 I(PSI)的协同工作。它通过裂解水产生 \ce {O2}、ATP 和 NADPH。
- 循环电子传递(Cyclic electron flow):仅涉及光系统 I(PSI)。它不产生 NADPH 和 \ce {O2},仅产生 ATP,用于调节细胞内 ATP 与 NADPH 的比例平衡。
这段课本内容通过三个核心实验,展示了人类如何从猜测走向实证,最终揭示光合作用本质的过程。我们可以结合大学生物化学的视角,对这些实验进行深度拆解:
希尔反应(1937 年):打破“二氧化碳分解释放氧气”的旧观念。在希尔(R. Hill)之前,人们普遍认为光合作用释放的氧气来自 \ce {CO2}。希尔最巧妙的贡献在于实现了 反应体系的简化与分离。
- 实验核心:他将离体的叶绿体放入盛有水和高铁盐(作为电子受体)的容器中,但不提供 \ce {CO2}。在光照下,他观察到了氧气的释放。
- 深度解读:
- 独立性:该实验证明了水的光解(光反应的一部分)可以不依赖于糖的合成(碳反应)而独立发生。
- 电子受体的本质:实验中的高铁盐(\ce {Fe^{3+}})起到了氧化剂的作用。在真实的叶绿体中,这个位置对应的是 \ce {NADP^{+}}。
- 回答讨论:希尔反应说明氧气可以在没有 \ce {CO2} 的情况下产生,暗示了水光解与糖合成是两个 相对独立 的过程,但直到鲁宾和卡门的实验才彻底证明了氧气中的氧元素 全部 来自水。
鲁宾和卡门实验(1941 年):氧气来源的“终极判决”。希尔反应虽然暗示了水是氧源,但并没有直接追踪氧原子的流向。鲁宾和卡门利用了当时最前沿的技术 ——同位素标记法。
- 实验逻辑:他们分别标记了 \ce {H2O} 和 \ce {CO2} 中的氧原子(使用 \ce {^{18} O})。
- 提供 \ce {H2O + C^{18} O2} → 释放的氧气是 \ce {O2}
- 提供 \ce {H2^{18} O + CO2} → 释放的氧气是 \ce {^{18} O2}
- 结论:这个实验提供了无可辩驳的直接证据,证明了光合作用释放的 \ce {O2} 全部来自 \ce {H2O}。
- 实验逻辑:他们分别标记了 \ce {H2O} 和 \ce {CO2} 中的氧原子(使用 \ce {^{18} O})。
阿尔农实验(1954、1957 年):发现能量的转化机器。如果说前两个实验搞清楚了“氧气从哪来”,阿尔农(D. Arnon)则搞清楚了“光能存到哪去”。
- 重大发现:他首先在光照下观察到叶绿体能合成 ATP(光合磷酸化)。随后他发现这一过程与水的光解是相伴发生的。
- 深度解读:这标志着人们认识到光能不仅仅是被用来“劈开”水分子,更被转换成了活跃的化学能储存在 ATP 和 NADPH 中。
总结:ATP 合成与希尔反应的关系。两者通过 电子传递链 耦合在一起:
- 光能 → 被色素捕获
- 希尔反应:水裂解产生电子(\ce {e^-})、质子(\ce {H^{+}})和 \ce {O2}
- 电子流:电子在传递链中流动释放能量,泵送 \ce {H^{+}} 形成跨膜梯度
- ATP 合成:质子梯度驱动 ATP 合酶,利用 ADP 和 Pi 合成 ATP
光合作用释放的氧气中的氧元素来自水,氧气的产生和糖类的合成不是同一个化学反应,而是分阶段进行的。实际上,光合作用的过程十分复杂,它包括一系列化学反应。根据是否需要光能,这些化学反应可以概括地分为 光反应(light reaction) 和 暗反应(dark reaction,现在也称为碳反应,carbon reaction) 两个阶段。
光反应阶段 是光合作用第一个阶段的化学反应,必须有光才能进行,这个阶段叫作光反应阶段。光反应阶段是在 类囊体的薄膜上 进行的。叶绿体中光合色素吸收的光能,有以下两方面用途:
一是将水分解为氧和 \ce {H^{+}},氧直接以氧分子的形式释放出去,\ce {H^{+}} 与氧化型辅酶 Ⅱ(\ce {NADP^{+}})结合,形成还原型辅酶 Ⅱ(\ce {NADPH})。\ce {NADPH} 作为活泼的还原剂,参与暗反应阶段的化学反应,同时也储存部分能量供暗反应阶段利用;
二是在有关酶的催化作用下,提供能量促使 ADP 与 Pi 反应形成 ATP。这样,光能就转化为储存在 ATP 中的化学能。这些 ATP 将参与第二个阶段合成有机物的化学反应。
水分解为氧和 \ce {H^{+}} 的同时,被叶绿体夺去两个电子。电子经传递,可用于 \ce {NADP^{+}} 与 \ce {H^{+}} 结合形成 \ce {NADPH}。
调压与保险机制。如果你想更全面地理解光反应,还可以补充两个细节:
- 调节机制(循环电子流):有时候植物不需要产生 \ce {NADPH},光能会被专门用来进行“循环电子传递”,此时光能只用于产生 ATP,而不产生 \ce {NADPH} 和 \ce {O2}。这通常是为了弥补卡尔文循环对 ATP 额外量的需求。
- 能量耗散(热能):并不是所有的光能都会被成功转化为化学能。当光照过强时,部分能量会以 热能 的形式耗散掉(非光化学猝灭),以保护叶绿体不被“烧坏”。
简单来说,课本抓住了“生产”的主干,而实际过程中还有“调压”和“保险”机制。
光系统
光系统是参与光合作用的蛋白质复合物的功能和结构单元,一起进行光合作用主要的光化学作用:光的吸收和转移能量和电子。光系统存在于植物、藻类和蓝细菌的类囊体膜中。类囊体位于植物和藻类的叶绿体中,以及光合细菌的细胞质膜中。存在两种光系统:光系统 II 和 光系统 I。
光系统的核心是 反应中心,它是一种利用光来还原分子(提供电子)的酶。该反应中心被光捕获复合物包围,以增强光的吸收。
在光系统中有两种反应中心:
- I 型反应中心(例如叶绿体中的光系统 I(P700)和绿硫细菌)
- II 型反应中心(例如叶绿体中的光系统 II(P680)和非硫紫色菌)
每个光系统可以通过其最具反应性的光的波长(对于叶绿体中的 PSI 和 PSII 分别为 700 和 680 纳米),光捕获复合物的数量和类型,以及所用的末端电子受体的类型来区分。
- I 型光系统 使用铁氧化物样铁 - 硫簇蛋白作为末端电子受体
- II 型光系统 最终将电子穿梭到醌末端电子受体
两种反应中心类型都存在于叶绿体和蓝细菌中,并且共同形成独特的光合链,能够从水中提取电子,从而产生作为副产物的氧气。
结构体:反应中心包含几(多于 24 或多于 33)个蛋白质亚基,其为一系列辅因子提供支架。辅因子可以是色素(如叶绿素、脱镁叶绿素、类胡萝卜素)、醌或铁 - 硫簇。
光系统 I 和 II 之间的关系:对于含氧光合作用,需要光系统 I 和光系统 II。有氧光合作用可以通过植物和蓝细菌进行;蓝细菌被认为是含有光系统的真核生物叶绿体的祖细胞。不能产生氧气的光合细菌具有与之相似的单一光系统。
当光系统 II 吸收光时,反应中心叶绿素中的电子被激发到更高的能级并被初级电子受体捕获。光激发的电子通过设置在类囊体膜中的电子传递链穿过 细胞色素 \ce {b6f} 蛋白复合体 到达光系统 I。利用这种能量降低(整个过程称为化学渗透),将氢离子(\ce {H^{+}})通过膜输送到类囊体腔中,以提供类囊体腔与叶绿体基质之间的潜在能量差异,相当于可用于产生 ATP 的质子动力。质子由质体醌运输。如果电子仅通过一次,则该过程称为非循环光合磷酸化。
当电子到达光系统 I 时,它填充光系统 I 的反应中心叶绿素的电子缺陷。当 ATP 合酶将存在于管腔中的质子通过膜转运到基质时产生 ATP。电子可以继续通过 PS I 周围的循环电子传递或通过铁氧还蛋白传递给酶 \ce {NADP^{+}} 还原酶。将电子和氢离子加入 \ce {NADP^{+}} 中以形成 \ce {NADPH}。这种还原剂被输送到卡尔文循环,与 3 - 磷酸甘油盐一起与 ATP 反应生成甘油醛 3 - 磷酸,这是植物可以制造各种物质的基本构件。
PSI 的进化起源:分子数据显示,PSI 可能是从绿色硫细菌的光系统发展而来。绿色硫细菌和蓝细菌、藻类和高等植物的光系统不一样,但是有许多类似的功能和类似的结构。不同光系统之间的三个主要特征相似:
- 氧化还原电位足以还原铁氧还蛋白
- 电子接受反应中心包括铁 - 硫蛋白
- 在两个光系统的复合物中的氧化还原中心构建在蛋白质亚基二聚体上
绿色硫细菌的光系统甚至包含 PSI 中电子传递链的所有相同辅助因子。两个光系统之间的相似性的数量和程度强烈地表明 PSI 来自绿色硫细菌的类似光系统。
光系统 II(PSII):光系统 II(或水 - 质体醌氧化还原酶)是氧光合作用的光依赖性反应中的第一个蛋白质复合物。它位于植物、藻类和蓝细菌的类囊体膜中。在光系统中,酶捕获光的光子以激发电子,然后通过各种辅酶和辅因子转移电子以将质体醌还原为质体酚。激发的电子氧化水而被取代,形成氢离子和分子氧。
通过光解水得到的电子来补充丢失的电子,光系统 II 为所有光合作用提供电子。由水的氧化产生的氢离子(质子)有助于产生质子梯度,ATP 合成酶用于产生 ATP。转移到质体醌的赋能电子最终用于还原 \ce {NADP^{+}} 为 \ce {NADPH} 或用于非循环电子流。
Z 字形(Z-scheme)路径 是描述非循环电子传递链中,电子能级随传递过程变化的示意图。之所以称为“Z 字形”,是因为电子流动的能级变化在坐标图(纵轴为氧化还原电位)上呈现出类似横放的“Z”字母形状。
这一过程详细分为以下四个核心阶段:
第一阶段:PSII 吸能与水的光解(Z 的第一画:向上升)
- 激发:光系统 II(PSII)中的 P680 叶绿素分子吸收光子,电子被激发到极高的能级并弹出。
- 补位:为了填补弹出电子留下的“空穴”,PSII 催化 水的光解(\ce {H2O -> O2 + H^{+} + e^{-}}),从水中夺取电子。这是地球大气中氧气的根本来源。
第二阶段:电子在“桥梁”中下坡传递(Z 的第二画:向下滑)
- 传递:电子从 PSII 弹出后,经由 质体醌(PQ)、细胞色素 \ce {b6f} 复合体 和 质体蓝素(Pc) 进行传递。
- 产能:电子在此过程中能级逐渐下降(“下坡”),释放出的能量被用于将 \ce {H^{+}} 从叶绿体基质泵入类囊体腔,形成质子梯度来驱动 ATP 的合成。
第三阶段:PSI 二次激发(Z 的第三画:再上升)
- 再加能:电子到达光系统 I(PSI)的 P700 时能级已较低。此时,PSI 吸收第二个光子,将电子再次提升到甚至比 PSII 还要高的能量水平。
第四阶段:生成 NADPH(Z 的第四画:最终下挫)
- 还原:高能电子经 铁氧还蛋白(Fd) 传递给 \ce {NADP^{+}} 还原酶。
- 终点:电子最终交给了 \ce {NADP^{+}},结合基质中的 \ce {H^{+}} 产生 \ce {NADPH}。
Z 字形路径完美解释了植物如何利用两次光能“接力”,将电子从能量极低的 水 提升到能量极高的 \ce {NADPH} 中,并在此过程中顺带生产了 ATP。
在大学生物学中,光反应的核心是 光合电子传递链。根据电子流动的路径不同,分为 非循环 和 循环 两种方式。
| 特性 | 非循环电子传递 (主流) | 循环电子传递 (调节) |
|---|---|---|
| 涉及光系统 | PSII 和 PSI | 仅 PSI |
| 电子来源 | 水 (\ce {H2O}) | PSI 自身循环 |
| 最终受体 | \ce {NADP^{+}} | 回到 PSI |
| 产物 | ATP、\ce {NADPH}、\ce {O2} | 仅 ATP |
| 主要功能 | 提供固碳所需的还原力和能量 | 补足 ATP 缺口,维持能量平衡 |
| 形象比喻 | “单行道”、“生产线” | “回力棒”、“跑圈”、“调压器” |
非循环电子传递 是光合作用最主要的路径,呈现为 Z 字形(Z-scheme)。电子从 水 出发,经过 PSII → 细胞色素 \ce {b6f} → PSI,最终交给 \ce {NADP^{+}},是“单行道”,不再回头。
循环电子传递 当细胞内 ATP 短缺或 \ce {NADPH} 过多时启动。电子从 PSI 弹出后传给 Fd,但 不流向 \ce {NADP^{+}},而是折返回细胞色素 \ce {b6f} 复合体,再回到 PSI,形成一个闭合循环 —— 就像“回力棒”或“在操场跑圈”。
简单来说,非循环流是“生产线”,而循环流是“调压器”。
光合磷酸化
在光合作用的过程中,以阳光作为能量源,将 ADP 磷酸化为 ATP 的过程称为 光合磷酸化。循环光磷酸化在有氧和无氧条件下都会发生。生命体只有两种能量来源:阳光和氧化还原反应。所有生物都会产生 ATP,它是生命的通用能源。在光合作用中,ATP 的合成通常包括水的光解或光解离,以及电子从水到光系统 II 的连续单向流动。
与氧化磷酸化的关联:光合磷酸化与呼吸作用中的氧化磷酸化原理极其相似,都利用 化学渗透(Chemiosmosis) 机制,即通过建立 \ce {H^{+}} 浓度梯度来驱动 ATP 合成酶。
区别:
- 氧化磷酸化:发生在线粒体,能量来自有机物氧化
- 光合磷酸化:发生在叶绿体类囊体膜,能量来自阳光
在光合磷酸化中,光能用于产生高能电子供体和低能电子受体。然后,电子通过电子传输链自发地从供体向受体移动。
ATP 是由一种称为 ATP 合酶 的酶合成的。在所有已知的生命形式中,该酶的结构及为其编码的基因都非常相似。卡尔文循环是光合作用最重要的部分。
ATP 合酶由跨膜电化学势梯度(通常以质子梯度的形式存在)驱动。电子传输链的功能是产生这种梯度。在所有生命体中,一系列氧化还原反应被用来产生跨膜的电化学势梯度或质子动力(pmf)。
循环光合磷酸化:这种形式的光合磷酸化作用发生在叶绿体的基质层或品格通道上。在循环光合磷酸化中,从 PS1 的 P700 释放的高能电子顺着一条环状路径流动。在循环电子流中,电子从称为光系统 I 的色素复合物开始,从主要受体传递至铁氧还蛋白,接着传递至质体醌,然后传递到细胞色素 \ce {b6f}(它类似于线粒体中发现的络合物),最后通过质体蓝素返回光系统 I。
该运输链产生质子动力,以将 \ce {H^{+}} 离子泵到膜外。这产生了一个浓度梯度,它在化学渗透过程中用来驱动 ATP 合酶。这条反应路径被称为循环光合磷酸化,它既不产生也不产生 \ce {NADPH}。与非循环光合磷酸化不同,电子没有被 \ce {NADP^{+}} 吸收,而是被送回细胞色素 \ce {b6f} 复合体。
细菌的光合作用中使用了一个光系统,因此它们也参与了循环光合磷酸化。厌氧条件下以及高辐照度和 \ce {CO2} 补偿点条件都对循环光合磷酸化有利。
非循环光合磷酸化:另一条反应途径是非循环光合磷酸化。它分为两个阶段,涉及两个不同的叶绿素光系统。非循环光合磷酸化发生在类囊体膜中,它是一个光反应。
首先,一个水分子被分解成,这个过程被称为光解(或光分裂)。其中的两个电子保留在光系统 II 中,而 \ce {2H^{+}} 和 \ce {O2} 则留作之后使用。然后,光系统反应核心中心周围的光合色素吸收一个光子。光激发这些色素中的电子,引起链式反应,最终将能量转移到光系统 II 的核心,并激发主要电子受体脱镁叶绿素的两个电子。电子的不足由另一个水分子中的电子来补充。
电子从脱镁叶绿素传输到质体醌,它从脱镁叶绿素接受 \ce {2e^{-}},从基质接受两个 \ce {H^{+}} 离子并合成。它后来被分解成 PQ,将两个电子释放到细胞色素 \ce {b6f} 复合,两个 \ce {H^{+}} 离子释放到类囊体腔中。接着,电子通过 Cyt \ce {b6} 和 Cyt \ce {f}。然后,它们被传递到质体蓝蛋白中,这个过程为氢离子(\ce {H^{+}})提供能量,以将其泵入类囊体空间。它产生了一个梯度,使 \ce {H^{+}} 离子流回到叶绿体的基质中,从而为 ATP 的再生提供能量。
光系统 II 复合物用外部的电子替代了损失的电子。但另外两个电子却没有像它们一样以类似循环路径的方式返回光系统 II。事实上,仍然被激发的电子被转移到一个光系统 I 络合物中,第二个来自太阳的光子将它们的能级提高到一个更高的水平。高激发电子被转移到一个受体分子上,这个受体被称为铁氧还蛋白 -\ce {NADP^{+}} 还原酶,该酶利用它们来催化反应:
\ce {NADP^{+} + 2H^{+} + 2e^{-} -> NADPH + H^{+}}
这个反应消耗了水分解产生的 \ce {H^{+}} 离子。最终,这个反应消耗了光子和水,并生成了 \ce {O2}、ATP、\ce {NADPH} 与 \ce {H^{+}}。
叶绿体中 \ce {NADPH} 的浓度高低可能有助于调节电子在光反应中经过哪条路径。当叶绿体为卡尔文循环提供的 ATP 数量较低时,\ce {NADPH} 会积累,使得反应路径可能会从非循环电子流转移到循环电子流。
为什么需要循环电子传递?这是一个非常敏锐的观察。在卡尔文循环中,消耗 ATP 和 \ce {NADPH} 的比例确实不是 1:1,而是 3:2。
简单来说,每固定 3 个 \ce {CO2} 分子,需要消耗 9 个 ATP 和 6 个 \ce {NADPH}。这种“不平衡”的原因在于卡尔文循环的三个阶段对能量的需求不同:
- 还原阶段(消耗 1:1):在将固定的碳(3 - PGA)还原为糖(G3P)的过程中,每处理一个碳需要 1 个 ATP 和 1 个 \ce {NADPH}。
- 再生阶段(额外消耗 ATP):为了让循环持续进行,植物必须把剩下的五分之六的糖(G3P)重新变回 \ce {CO2} 的受体(RuBP)。这个 五碳糖的再生过程 只需要 3 个额外的 ATP,而不需要 \ce {NADPH}。
这就是为什么 ATP 的需求量更大。这也解释了我们之前提到的 循环电子传递链 存在的意义:它能让植物在不产生 \ce {NADPH} 的情况下,单独“加餐”生产 ATP,以满足这个 3:2 的比例缺口。
植物何时开启循环流?:这种“加餐”行为通常发生在植物需要 打破 ATP 和 \ce {NADPH} 生产与消耗的失衡 时:
补足卡尔文循环的 ATP 缺口:非循环电子流产生的 ATP 和 \ce {NADPH} 比例大约是 1:1,但卡尔文循环固定 \ce {CO2} 时消耗两者的比例是 3:2。为了填补这 0.5 份 ATP 的差额,植物必须开启循环流来单独生产 ATP。
\ce {NADPH} 积累(还原力过剩):当细胞内 \ce {NADP^{+}} 含量低、\ce {NADPH} 堆积时,电子没有足够的受体去生成更多的 \ce {NADPH}。此时电子会“折返”进行循环流动,只用来建立质子梯度产生 ATP,从而调节两者的比例。
特定的固碳策略(如 C4 植物):C4 植物的维管束鞘细胞通常缺乏 PSII 活性,主要运行循环电子流。这是因为它们需要大量的额外 ATP 来驱动 \ce {CO2} 浓缩机制,同时要尽量减少 \ce {O2} 的产生以降低光呼吸。
环境胁迫响应:在强光、干旱等逆境下,植物可能通过增加循环流来产生额外的 ATP,用于修复受损的蛋白质或维持渗透压平衡,同时避免产生过多的还原力导致氧化损伤。
简单来说,当植物发现“钱(ATP)不够花”或者“原料(\ce {NADPH})囤积太多”时,就会启动循环流这个“调压器”。
非循环电子传递链
非循环电子传递(Non-cyclic Electron Flow)是光合作用最主要的路径,呈现为“Z”字形(Z-scheme)。
路径:\ce {H2O ->} 光系统 II (PSII) \ce {->} 细胞色素 \ce {b6f} 复合体 \ce {->} 光系统 I (PSI) \ce {-> \ce {NADP^{+}}}
详细步骤:
水的光解:光能激发 PSII 的反应中心(P680),电子被弹出。为了填补空缺,PSII 氧化水分子,释放出 \ce {O2} 和 \ce {H^{+}}。
质子泵送:电子经质体醌(PQ)传递给细胞色素 \ce {b6f} 复合体。在此过程中,能量被用来将 \ce {H^{+}} 从叶绿体基质泵入类囊体腔,形成质子梯度用于合成 ATP。
二次激发:电子经质体蓝素(PC)传至 PSI。光能再次激发 PSI 反应中心(P700),电子获得更高能量。
生成 \ce {NADPH}:高能电子最终经铁氧还蛋白(Fd)传递给 \ce {NADP^{+}},生成 \ce {NADPH}。
产物:ATP、\ce {NADPH} 和 \ce {O2}。
Z 字形路径完美解释了植物如何利用两次光能“接力”,将电子从能量极低的 水 提升到能量极高的 \ce {NADPH} 中,并在此过程中顺带生产了 ATP。
这个过程产生的 ATP 和 \ce {NADPH} 会被送往 卡尔文循环。
循环电子传递链
循环电子传递(Cyclic Electron Flow)当细胞内 ATP 短缺或 \ce {NADPH} 过多时,植物会启动这种“旁路”来调节能量比例。
路径:仅涉及光系统 I (PSI) 和细胞色素 \ce {b6f} 复合体。
过程:
电子从 PSI 弹出后传给 Fd,但 不流向 \ce {NADP^{+}},而是折返回细胞色素 \ce {b6f} 复合体。
电子再次通过复合体流回 PSI,形成一个闭合循环。
只泵质子,不还原:这个循环不断重复泵送 \ce {H^{+}} 的过程,建立质子梯度来合成 ATP。
特点:不产生 \ce {NADPH},也不产生 \ce {O2}(因为不涉及 PSII 和水的光解)。
产物:仅产生 ATP。
之所以被称为“循环”,是因为电子在光系统 I(PSI)中形成了一个 闭合回路。电子从 PSI 的反应中心弹出后,传递给铁氧还蛋白(Fd),但它并不流向 \ce {NADPH},而是被回收到细胞色素 \ce {b6f} 复合体中。电子随后经由质体蓝素(Pc)再次回到 PSI,就像在操场跑圈一样周而复始。
这个过程不涉及 PSII,所以 不裂解水,也 不产生 \ce {O2} 和 \ce {NADPH}。它唯一的功劳是利用电子流动产生的能量泵送质子,从而只驱动 ATP 的合成。
NADH 与 NADPH 的区分
区分这两者其实有一个非常简单的“字母记忆法”:
如何区分光合还是呼吸?
看字母 P:
- \ce {NADPH}:含有字母 P,对应 Photosynthesis(光合作用)。它是光反应产生的还原力,专门用于暗反应(卡尔文循环)中合成糖类等有机物(合成代谢)。
- \ce {NADH}:没有字母 P,对应呼吸作用(Cellular Respiration)。它在糖酵解和三羧酸循环中产生,最后去线粒体内膜“发电”产生 ATP(分解代谢)。
如何记忆辅酶 I 和辅酶 II?
- 辅酶 I(Coenzyme I):对应 \ce {NAD^{+}}
- 辅酶 II(Coenzyme II):对应 \ce {NADP^{+}}
记忆窍门:辅酶 II 比辅酶 I 多了一个磷酸基团(Phosphate)。既然“II”比“I”大,那么含有“额外赠品(P)”的就是辅酶 II。
为什么课本把 \ce {NADH} 记作 [\ce {H}]?
这主要是高中阶段的简化处理。
- 在呼吸作用中,\ce {NADH} 和 \ce {FADH2} 都携带氢原子(质子和电子),为了方便理解,课本统称它们为 还原型辅酶 [\ce {H}]。
- 但在大学生物学中,必须区分开:\ce {NADH} 主要由脱氢酶产生,而 \ce {NADPH} 是光合作用特有的高能还原态载体,两者不能混用。
核心区别总结:
| 特性 | \ce {NADH} (辅酶 I) | \ce {NADPH} (辅酶 II) |
|---|---|---|
| 字母 P | 无(单纯的载体) | Phosphate(磷酸基团) |
| 主要领域 | 呼吸作用(分解代谢) | 光合作用(合成代谢) |
| 细胞比率 | \ce {NAD^{+}} / \ce {NADH} 比例高(利于吸电子氧化) | \ce {NADP^{+}} / \ce {NADPH} 比例低(利于供电子还原) |
简单来说,\ce {NADH} 是为了 “拆解”食物换钱(ATP),而 \ce {NADPH} 是为了 “建设”(合成大分子)。这种“分工明确”的机制让细胞能独立调节合成与分解过程,互不干扰。
暗反应阶段
暗反应概述
一般来说,暗反应阶段(Dark Reaction Stage)是光合作用第二个阶段中的化学反应,它 不直接依赖光,在叶绿体的基质中进行。在这一阶段,\ce {CO2} 被利用,经过一系列的反应后生成糖类。
应当说明,暗反应名称并不严谨,叫作碳反应更好。 因为这些反应虽然不直接需光,但需要光反应提供的 ATP 和 \ce {NADPH} 产物,且部分酶也受光调节。
\ce {CO2} 是如何转变成糖类的呢?20 世纪 40 年代,美国科学家卡尔文(M. Calvin, 1911—1997)等用小球藻(一种单细胞的绿藻)做了这样的实验:用经过 \ce {^{14} C} 标记的 \ce {^{14} CO2},供小球藻进行光合作用,然后追踪放射性 \ce {^{14} C} 的去向,最终探明了 \ce {CO2} 中的碳是如何转化为有机物中的碳的。
绿叶通过气孔从外界吸收的 \ce {CO2},在特定酶的作用下,与 C5(一种五碳化合物)结合,这个过程称作 \ce {CO2} 的固定。一分子的 \ce {CO2} 被固定后,很快形成两个 C3 分子。在有关酶的催化作用下,C3 接受 ATP 和 \ce {NADPH} 释放的能量,并且被 \ce {NADPH} 还原。随后,一些接受能量并被还原的 C3,在酶的作用下经过一系列的反应转化为糖类;另一些接受能量并被还原的 C3,经过一系列变化,又形成 C5。这些 C5 又可以参与 \ce {CO2} 的固定。这样,暗反应阶段就形成从 C5 到 C3 再到 C5 的循环,可以源源不断地进行下去,因此暗反应过程也称作 卡尔文循环。
简而言之,在光反应阶段,光能被叶绿体内类囊体膜上的色素捕获后,将水分解为 \ce {O2} 和 \ce {H^{+}} 等,形成 ATP 和 \ce {NADPH},于是光能就转化成 ATP 和 \ce {NADPH} 中的化学能;ATP 和 \ce {NADPH} 驱动在叶绿体基质中进行的暗反应,将 \ce {CO2} 转化为储存化学能的糖类。可见光反应和暗反应紧密联系,能量转化与物质变化密不可分。光合作用产生的有机物,不仅供植物体自身利用,还养活了包括你我在内的所有异养生物。光能通过驱动光合作用而驱动生命世界的运转。
在大学生物学中,高中所学的“暗反应”现在学术界更准确地称其为 固碳反应(Carbon fixation) 或 碳反应(Carbon reaction),因为这些反应虽然不直接需光,但需要光反应提供的 ATP 和 \ce {NADPH} 产物,且部分酶也受光调节。
它根据途径不同细分为:
- 卡尔文循环(Calvin cycle):也称光合碳还原循环(C3 途径)。这是所有植物固碳的核心途径,直接产物是三碳糖(G3P)。
- C4 类二氧化碳固定(C4 pathway):主要见于玉米等热带植物。它通过在叶肉细胞中将 \ce {CO2} 固定为四碳酸,再转运至维管束鞘细胞释放,从而克服“光呼吸”。
- 景天酸代谢(CAM):见于仙人掌等干旱植物。这类植物在夜间固定 \ce {CO2} 形成有机酸,白天再释放 \ce {CO2} 进入卡尔文循环,以最大限度减少水分散失。
总结:高中的“光反应”在大学里深入到了 电子传递链 的能级变化,而“暗反应”则扩展为植物对不同环境适应的 多元固碳策略。
这三种植物由于 固碳途径 的差异,在不同的环境下展现出截然不同的生存智慧和效率:
| 类型 | 策略 | 代表植物 | 适应环境 | 光呼吸 |
|---|---|---|---|---|
| C3 | 直接卡尔文循环 | 水稻、小麦 | 温带、湿润 | 高损耗(约 30%) |
| C4 | 空间分离(\ce {CO2} 泵) | 玉米、甘蔗 | 热带高温 | 几乎为零 |
| CAM | 时间分离(昼夜节律) | 仙人掌、凤梨 | 极端干旱 | 极低 |
- C3 植物(温带代表):二氧化碳固定最初产物是 3 - 磷酸甘油酸(C3)。在温和湿润环境下效率尚可,但在高温下,气孔关闭会导致 \ce {CO2} 不足,面临严重的 光呼吸 损耗。
- C4 植物(高温强者):通过 空间分离(叶肉细胞固定,\ce {CO2} 泵往维管束鞘细胞再次固定)来浓缩 \ce {CO2}。这使得它们在强光和高温下固碳效率极高,且几乎没有光呼吸。
- CAM 植物(干旱专家):采取 时间分离。夜间开启气孔吸收 \ce {CO2} 并储存在有机酸中,白天关闭气孔以节水,再释放 \ce {CO2} 供光合作用。虽然节水能力最强,但其光合速率和生长速度通常较慢。
总结:C4 是 “空间效率派”,CAM 是 “极限节水派”,而 C3 则是 “中庸派”。
卡尔文循环
卡尔文循环(英语:Calvin cycle,或简称卡氏循环,又译作开尔文循环,或光不依赖反应、生物合成相、暗反应)是由美国加州大学伯克利分校梅尔文・卡尔文、安德鲁・本森(Andrew Benson)和詹姆斯・巴沙姆(James Bassham)3 人发现。梅尔文・卡尔文于 1961 年获得诺贝尔化学奖。
卡尔文循环是光合作用里碳反应的一部分,反应场所为叶绿体内的基质,分为三个阶段:羰化、还原和二磷酸核酮糖的再生。
卡尔文循环是一种类似克雷伯氏循环的新陈代谢过程,其可使起始物质以分子的形态进入和离开这循环后发生再生。碳以二氧化碳形态进入,并以糖的形态离开。整个循环是利用 ATP 作为能量来源,并以降低能阶的方式来消耗 \ce {NADPH},如此以增加高能电子来制造糖。其制造出来的碳水化合物并不是葡萄糖,而是一种称为 3 - 磷酸甘油醛的三碳糖。为了要合成 1 摩尔这种碳,整个循环过程必须发生 3 次的取代作用,将 3 摩尔的二氧化碳固定。
尽管它也被称为“暗反应”,但卡尔文循环实际上并不发生在黑暗或夜间。这是因为该过程需要 \ce {NADPH},而 \ce {NADPH} 的寿命很短,并且来自光依赖反应。在黑暗中,植物将淀粉储备中的蔗糖释放到韧皮部,为植物提供能量。因此,当光照独立于光合作用类型(C3 类二氧化碳固定、C4 类二氧化碳固定和景天酸代谢 (CAM))时,就会发生卡尔文循环;景天酸代谢植物每天晚上将苹果酸储存在液泡中,并在白天释放出来,以使这一过程发挥作用。
卡尔文循环分为三个阶段:
- 羰化阶段(\ce {CO2} 固定):\ce {CO2} 与 C5(1,5 - 二磷酸核酮糖,RuBP)结合,形成不稳定的六碳化合物,随即分解为两分子的三碳化合物(3 - 磷酸甘油酸,3 - PGA)。
- 还原阶段(消耗 1:1):在将固定的碳(3 - PGA)还原为糖(G3P)的过程中,每处理一个碳需要 1 个 ATP 和 1 个 \ce {NADPH}。
- 再生阶段(额外消耗 ATP):为了让循环持续进行,植物必须把剩下的五分之六的糖(G3P)重新变回 \ce {CO2} 的受体(RuBP)。这个 五碳糖的再生过程 只需要 3 个额外的 ATP,而不需要 \ce {NADPH}。

为什么需要额外的 ATP?在卡尔文循环中,消耗 ATP 和 \ce {NADPH} 的比例确实不是 1:1,而是 3:2。
简单来说,每固定 3 个 \ce {CO2} 分子,需要消耗 9 个 ATP 和 6 个 \ce {NADPH}。这种“不平衡”的原因就在于上述的第三阶段 ——五碳糖的再生过程只需要 3 个额外的 ATP,而不需要 \ce {NADPH}。
这就是为什么 ATP 的需求量更大。这也解释了为什么植物需要 循环电子传递链:它能让植物在不产生 \ce {NADPH} 的情况下,单独“加餐”生产 ATP,以满足这个 3:2 的比例缺口。
光呼吸
光呼吸(英语:photorespiration)是所有使用卡尔文循环进行碳固定的细胞在光照和高氧低二氧化碳情况下发生的一个生化过程。它是卡尔文循环中一个损耗能量的副反应。过程中氧气被消耗,并且会生成二氧化碳。如果光呼吸发生在进行光合作用的生物中,那么光呼吸会抵消约 30% 的光合作用。因此降低光呼吸被认为是提高光合作用效能的途径之一。但是人们后来发现,光呼吸有着很重要的细胞保护作用。
在光呼吸过程中,参与卡尔文循环的反应物 1,5 - 二磷酸核酮糖(英文缩写为 RuBP)和催化剂核酮糖 - 1,5 - 二磷酸羰化酶/加氧酶(英文缩写为 RuBisCO)发生了与其在光合作用中不同的反应。光合作用中,二磷酸核酮糖在羰化/加氧酶的催化下与二氧化碳结合增加一个碳原子,再经过一系列反应,最终生成 3 - 磷酸甘油酸。后者再经过部分卡尔文循环中的步骤,可再次重新生成为二磷酸核酮糖。但光呼吸过程中,二磷酸核酮糖在羰化/加氧酶的催化下生成 2 - 磷酸乙醇酸。
换而言之,在羰化/加氧酶的作用下,二磷酸核酮糖参与了两种过程:生成能量获得碳素的卡尔文循环,以及消耗能量释放碳素的光呼吸。由此可见,光呼吸和卡尔文循环关系密切,它们之间的关系可以作一形象的理解:糖工厂内(行卡尔文循环的细胞)的葡萍糖生产线(卡尔文循环)因一部机器(1,5 - 二磷酸核酮糖羰化酶/加氧酶)构造不完善,一部分原材料(1,5 - 二磷酸核酮糖)不断被错误加工,产出次品(2 - 磷酸乙醇酸),虽然有一补救措施,可将次品重加工并再次投入生产线,但是整个过程却是非常费时费力的。这个错误加工和补救的过程就是光呼吸。
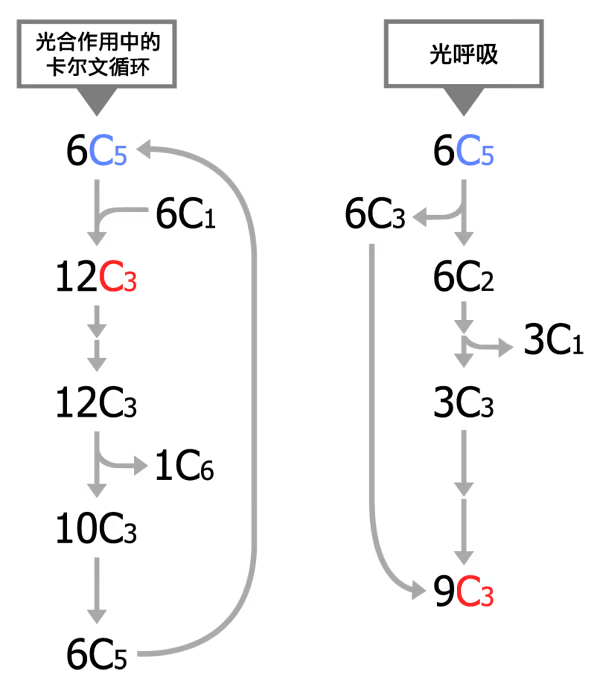
在 \ce {O2} 浓度过高或 \ce {CO2} 浓度过低的情况下(例如炎热干旱导致气孔关闭时),催化固碳的 Rubisco 酶 会表现出“两面性”:它既能催化 \ce {CO2} 的固定(羰化酶活性),也能催化 C5(RuBP,五碳化合物)与 \ce {O2} 反应(加氧酶活性)。
这个过程之所以被视为“浪费”,主要有以下几个原因:
- 消耗 C5:Rubisco 错误地让 \ce {O2} 氧化了原本用于循环的 C5,而不是将其用于固定 \ce {CO2}。
- 释放 \ce {CO2}:反应产生的二碳化合物经过复杂的代谢路径后,最终会释放出 \ce {CO2},把好不容易固定下来的碳又丢掉了。
- 不产生能量:与细胞呼吸不同,光呼吸过程不仅 不产生 ATP,反而还要 消耗 ATP。
这就是为什么 C4 植物(如玉米)进化出了特殊的结构来浓缩 \ce {CO2},从而规避这种由于 \ce {O2}“竞争”导致的能量浪费。

虽然光呼吸会消耗能量且流失 \ce {CO2},但在进化中被保留下来,说明它具有重要的生理意义:
- 保护作用:在强光、干旱等逆境下(气孔关闭时),光呼吸可以消耗过剩的光能和还原力,防止 光合机构被“烧坏”。
- 代谢调节:它有助于消除有毒的中间产物,并参与氮代谢的补充。
所有这些发现导致了植物科学方面的讨论,是否应该在降低植物光呼吸方面去作出努力。
C4 类二氧化碳固定
在 15 亿年前,随着光合作用的出现,氧气开始在地球的大气层积累。二氧化碳固定过程中的关键酶 RuBisCO 同时具有加氧酶的功能,它在一个重要的副反应里也催化了氧的固定。氧气可以与二氧化碳竞争 RuBisCO 的活性部位。在原始大气里,氧气缺乏,在上面提到的副反应里面,二碳化合物积累,碳循环受阻,同化作用在这种环境下并不能顺利进行。回收二碳化合物的过程对于植物来说也是费时耗力的。此过程需要耗氧,人们称之为光呼吸。
随着温度的升高,RuBisCO 与氧气的亲和力递增迅速,超过了对二氧化碳的递增速度,这对于生长在干旱热带地区的植物来说并不是好消息,它们需要另外的途径以固定二氧化碳。植物发展出“ATP 驱动的 \ce {CO2} 泵”,从而创造出一种与原始大气相似的内环境。
除了 Rubisco - 反应外,叶肉细胞还发展出 PEP - 羰化途径以固定二氧化碳。在这个过程里 \ce {CO2} 会被磷酸烯醇式丙酮酸(缩写 PEP)所固定,之后生成四碳化合物草酰乙酸(缩写 OAA),这就是 C4 类植物名称的由来。草酰乙酸转换为苹果酸或天门冬氨酸后进入维管束鞘,在苹果酸酶的作用下生成丙酮酸(Pyruvat)和 \ce {CO2}。在维管束鞘里 \ce {CO2} 浓度高,卡尔文循环能高效的运行。
1960 年代,澳洲科学家哈奇(M. D. Hatch)和斯莱克(C. R. Slack)发现玉米、甘蔗等热带绿色植物,除了和其他绿色植物一样,具有卡尔文循环外,\ce {CO2} 首先通过一条特别的途径被固定。这条途径也被称为 “哈奇 - 史莱克途径”。
C4 植物主要是一些生活在干旱热带地区的植物。在这种环境中,植物倘若长时间开放气孔吸收二氧化碳,会导致水分通过蒸腾作用过快的流失。所以,植物只能短时间开放气孔,二氧化碳摄入量必然少。植物必须利用这少量的二氧化碳进行光合作用,合成自身所需物质。
在 C4 植物叶片维管束的周围,有维管束鞘包围,这些维管束鞘细胞含有叶绿体,但里面并无基粒或发育不良。在这里,就是主要进行卡尔文循环的地方。
其叶肉细胞中,含有独特的酶,即磷酸烯醇式丙酮酸羰基化酶,使得二氧化碳先被一种三碳化合物磷酸烯醇式丙酮酸同化,形成四碳化合物草酰乙酸,这也是该暗反应类型名称的由来。这草酰乙酸在转变为苹果酸盐后,进入维管束鞘,就会分解释放二氧化碳和一分子丙酮酸。二氧化碳利用此细胞才进入卡尔文循环,后同 C3 进程。而丙酮酸则会被再次合成磷酸烯醇式丙酮酸。此过程消耗 ATP。
该类型的优点是,二氧化碳固定效率比 C3 高很多,有利植物在干旱环境生长。C3 植物行光合作用所得的淀粉,会贮存在叶肉细胞中,因为这是卡尔文循环的场所,而维管束鞘细胞则不含叶绿体。而 C4 植物的淀粉,将会贮存于维管束鞘细胞 (bundle-sheath cells) 内,因为 C4 植物的卡尔文循环是在此发生的。
20 世纪 60 年代,马沙・哈奇和罗杰・斯莱克阐明了这种发生在相邻两种类型细胞里的四碳双羰酸途径的反应,后世便以他们的名字命名该循环。循环开始于叶肉细胞,但那里缺少 RuBisCO,反应转到维管束鞘里面进行,在这里,就遵循 C3 类植物的卡尔文循环途径发生反应。
C4 植物的维管束鞘细胞通常缺乏 PSII 活性,主要运行 循环电子流。这是因为它们需要大量的额外 ATP 来驱动 \ce {CO2} 浓缩机制,同时要尽量减少 \ce {O2} 的产生以降低光呼吸。
简单来说,当植物发现“钱(ATP)不够花”或者“原料(\ce {NADPH})囤积太多”时,就会启动循环流这个“调压器”。而 C4 植物由于需要额外的 ATP 来驱动其独特的 \ce {CO2} 泵机制,对循环电子流的依赖更强。
景天酸代谢
景天酸代谢(英语:Crassulacean acid metabolism,简称 CAM,也称为 CAM 光合作用)是一种特殊而精巧的碳固定方式,它在某些植物中进化,以适应干旱条件。这一代谢途径最早在景天科植物中被发现,由此得名。代表性的植物有仙人掌、凤梨和长寿花等,99% 的仙人掌物种都采用景天酸代谢。
大部分植物开放气孔吸收二氧化碳,二氧化碳参与一系列光合反应生成生命活动所需的有机物。然而,气孔的开放伴随着水分蒸发流失,对于生长在炎热干旱地区的植物非常不利。因此,CAM 植物演化出一套生存机制,只在凉爽的夜间开放气孔,将二氧化碳以有机酸的形式暂时储存下来,到白天再进行卡尔文循环,此时可以闭合大部分气孔,有效减少水分流失。
CAM 固碳途径与 C4 固碳途径有一定的相似之处,两者利用相似的化学物质作为媒介,都通过拆分碳固定的步骤,实现对干旱环境的适应。相比之下,C4 类植物实行的是 空间分离(叶肉细胞和维管束鞘细胞两个相对独立的场所),而景天酸代谢植物则服从昼夜节律,细节如下:
夜间:二氧化碳进入细胞质,在磷酸烯醇式丙酮酸(PEP)的作用下生成草酰乙酸(OAA),后续被还原为苹果酸,并储存于细胞的液泡中。PEP 羰化酶催化草酰乙酸的生成,该酶的表达受高温(即白天)和苹果酸的抑制。虽然吸收了二氧化碳,但将其转化为有机物的过程需要光反应产物的参与,因此卡尔文循环无法继续进行。
白天:液泡中的酸性物质(主要是苹果酸,但也有天门冬氨酸)会被脱羰,释放的二氧化碳进入叶绿体,参与卡尔文循环。
景天酸代谢植物必须准备足够的 PEP 以供夜间二氧化碳固定使用。为此植物在日间储存淀粉,晚间它们将通过丙酮酸转变为磷酸烯醇式丙酮酸。
大多数景天酸代谢植物的生态特征可大致归为如下三类:
沙漠及半干旱地区的陆生植物:西半球沙漠生境中的代表性景天酸代谢类群包括仙人掌科、天门冬科(龙舌兰亚科)等,而东半球的类似生境中则有大载科、番杏科、阿福花科(芦荟属)等。开启气孔会导致水分蒸腾流失,因而干旱环境会限制植物进行气体交换,而气孔“夜开日合”的景天酸代谢模式则可以帮助植物保存体内水分,一些适应干旱的景天酸代谢物种在数周甚至数月不进行呼吸作用的情况下依然可以存活,由此导致的较低的光合作用速率也是许多沙漠植物类群生长缓慢的原因。
热带及亚热带森林地区的附生植物:近一半的景天酸代谢植物都原生于降水量中等或较高的地区,例如巴拿马和马达加斯加雨林中的一些兰科和凤梨科物种。这些相对矮小的附生植物依附在高大乔木的树干或枝条上,以在树冠层叠的丛林环境中获得相比地面更多的阳光;这样的生长策略使得它们无法从土壤中获取水分,而景天酸代谢可以帮助它们在水分受限的条件下提高用水效率。一些兰花的气生根也可进行景天酸代谢。
水生植物:尽管水对于水生植物并不是稀缺资源,部分水生植物却演化出了景天酸代谢的表型,且在维管植物的多个大类群中都可以找到独立演化的代表类群(石松类的水乞科、单子叶植物的慈姜属、真双子叶植物中的水生青锁龙 Crassula aquatica 等)。这些物种多分布于寡营养湖泊或季节性浅水塘,生境中的二氧化碳水平常有较大波动。一种较广接受的观点是采用景天酸代谢可帮助植物体应对水中较低的二氧化碳含量。
藻类和细菌
真核藻类,如红藻、绿藻、褐藻等,和植物一样具有叶绿体,也能够进行产氧光合作用。光被叶绿素吸收,而很多藻类的叶绿体中还具有其它不同的色素,赋予了它们不同的颜色。
进行光合作用的细菌不具有叶绿体,而直接由细胞本身进行。属于原核生物的蓝细菌(或者称“蓝藻”)同样含有叶绿素,和叶绿体一样进行产氧光合作用。事实上,目前普遍认为叶绿体是由蓝细菌演化而来的。其它光合细菌具有多种多样的色素,称作细菌叶绿素或菌绿素,但不氧化水生成氧气,而以其它物质(如硫化氢、硫或氢气)作为电子供体。不产氧光合细菌包括紫硫细菌、紫非硫细菌、绿硫细菌、绿非硫细菌和太阳杆菌(Heliobacteria)等。